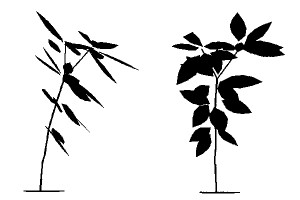
| 恾侾丂 寁嶼婡撪偱嵞峔惉偟偨椦彴偺傾僆僉偺3師尦峔憿丅 乮嵍乯 恾偺岦偐偭偰塃懁偵摴楬偑偁偭偰岝偑幩偟崬傓丅乮塃乯 摨偠屄懱傪摴楬偺懁偐傜尒偨偲偙傠丅 |
僥儗價偺惗偒暔斣慻偺戣嵽偼丄傎偲傫偳偡傋偰偑摦暔偩丅偨偟偐偵摦暔偺峴摦偼尒偰偄偰朞偒側偄丅 恖娫偵廳偹崌傢偣偰尒傞偙偲偑偱偒傞偟丄偦偺偄偐偵傕尗偄傆傞傑偄偵姶怱偡傞偙偲傕懡偄丅偄偭傐偆丄怉暔偑戣嵽偵側傞偺偼傕偭傁傜墍寍斣慻偽偐傝偱偁傞丅怉暔偺惗偒曽偺偍傕偟傠偝傪僥乕儅偵偟偨斣慻側偳傔偭偨偵尒傞偙偲偑偱偒側偄丅
偱傕丄摦暔偑尗偄偲偄偆偺側傜怉暔偩偭偰尗偄丅摦暔偺峴摦偺側偐偵棟偵偐側偭偨"崌棟惈"偑尒弌偝傟傞偺偲摨條偵丄怉暔偺惗偒曽偵傕懡偔偺崌棟惈傪尒弌偡偙偲偑偱偒傞丅怉暔偵偍偄偰傕摦暔偵偍偄偰傕丄挿偄恑壔偺楌巎偺側偐偱惗懚偲斏怋偵傛傝揔偟偨惗偒曽偑慖戰偝傟偰偒偨偙偲偵堘偄偼側偄丅
偦偺傛偆側崌棟惈偼嵶朎偺拞側偳儈僋儘偺悽奅偵傕偨偔偝傫媗傑偭偰偄傞偑丄栚偱尒偊傞儅僋儘側峔憿偺拞偵傕傂偦傫偱偄傞丅摦暔偺傛偆偵摦偒夞偭偰僄僒傪庢傞偙偲偼側偄偐傢傝偵丄怉暔偺帒尮妉摼偱偼偦偺俁師尦揑側峔憿偑壥偨偡栶妱偑偲偰傕戝偒偄丅僄僱儖僊乕尮偱偁傞岝傪妉摼偡傞偵偼抧忋晹偺嬻娫峔憿偑丄傑偨搚偺拞偺悈傗塰梴暘傪廤傔傞偵偼崻偺嬻娫峔憿偑僇僊偲側傞丅偄偭偰傒傟偽丄懡偔偺摦暔偑峴摦偱彑晧偟偰偄傞傛偆偵丄怉暔偼儅僋儘側峔憿偡側傢偪'宍'偱彑晧偟偰偄傞丅偁傜偨側嬻娫傊偲巬傗崻傪怢偽偟偰帒尮傪妉摼偟丄偦偺堦曽偱埲慜偵嶌偭偨婍姱傪屚傜偟偰偄偔偝傑傪丄怉暔偺'峴摦'偲屇傇恖傕偄傞丅
杮峞偱偼丄堦枃堦枃偺梩偺岦偒丄堦杮偺宻傊偺梩偺晅偒偐偨丄栘偺巬暘偐傟丄偦偟偰巬偺怢挿丒屚巰偵傛傞僟僀僫儈僢僋側庽宍宍惉傪偲傝偁偘偰丄怉暔偺宍亖儅僋儘側峔憿偑岝妉摼婡擻偲偳偺傛偆偵娭傢偭偰偄傞偺偐傪徯夘偡傞丅
梩偼岝傪庴偗偰岝崌惉傪偡傞婍姱偱偁傞丅傛傝懡偔偺岝傪庴偗傟偽傛傝懡偔偺岝崌惉嶻暔丄偡側傢偪桳婡暔傪嶌傞偙偲偑偱偒傞丅岝崌惉偵傛傞桳婡暔偺嫙媼側偟偵偼丄怉暔偼惉挿偡傞偙偲傕惗偒偰偄偔偙偲傕偱偒側偄丅椦傗憪傓傜偺撪晹偺傛偆偵岝僄僱儖僊乕偑晄懌偟偑偪側娐嫬偱偼丄側傞傋偔偨偔偝傫岝傪庴偗傜傟傞傎偆偑怉暔偵偲偭偰偼偁傝偑偨偄丅岝偑偳偺曽岦偐傜傕嬒堦偵棃傞側傜偽梩偑偳偪傜傪岦偄偰偄傛偆偲庴偗傞岝偺検偵偼娭學側偄偑丄岝尮偵曽岦惈偑偁傞偲偒偵偼丄岝偑偍傕偵棃傞曽岦偵懳偟偰悅捈偵梩傪傂傠偘偨傎偆偑庴岝検傪戝偒偔偡傞偙偲偑偱偒傞丅
偨偟偐偵丄椦偺墢嬤偔偺怉暔偱偼丄椦偺奜偺曽岦偵庴岝柺傪岦偗傞傛偆偵孹偄偨梩偑偟偽偟偽尒傜傟傞丅傑偨丄椦偺側偐偱栘偑屚傟偰偱偒偨寗娫偺嬤偔偱傕丄偦偺寗娫偺曽岦偵岦偐偭偰孹偄偨梩傪懡偔尒傞丅偄偐偵傕擔摉偨傝偑傛偄傛偆偵梩偺岦偒傪挷惍偟傑偟偨丄偲偄偆報徾偱偁傞丅
偙偆偟偨偐偨偪偑杮摉偵庴岝検傪戝偒偔偡傞偺偵栶棫偭偰偄傞偺偐傪妋偐傔傞偨傔偵丄椦偺壓偱惗堢偟偰偄傞忢椢掅栘偺傾僆僉偺俁師尦峔憿傪應掕偟偨椺傪徯夘偟傛偆(1)丅屄乆偺梩偺嬻娫攝抲偺僨乕僞傪巊偭偰傾僆僉偺宍傪寁嶼婡偺拞偵嵞尰偟(恾1)丄偝傜偵嬻偺偳偪傜偺曽岦偑偳偺偔傜偄柧傞偄偐丄忋傪暍偭偰偄傞攚偺崅偄怉暔偵偳偺偖傜偄擔堿偵偝傟偰偄傞偐丄偲偄偆忣曬偲慻傒崌傢偣傞偲丄偦傟偧傟偺梩偑庴偗傞岝偺検傪寁嶼偡傞偙偲偑偱偒傞(2)丅
|
| 恾侾丂 寁嶼婡撪偱嵞峔惉偟偨椦彴偺傾僆僉偺3師尦峔憿丅 乮嵍乯 恾偺岦偐偭偰塃懁偵摴楬偑偁偭偰岝偑幩偟崬傓丅乮塃乯 摨偠屄懱傪摴楬偺懁偐傜尒偨偲偙傠丅 |
忋憌偺栘偺栁傝偐偨傗椦偺墢偺塭嬁側偳偵傛傝丄岝偺棃傞曽岦偵偼偐偨傛傝偑偁傞丅寁嶼偺寢壥丄傾僆僉偺屄乆偺梩偺岦偒偼偦偺応強偱傕偭偲傕懡偔岝傪庴偗傜傟傞曽岦偲傎傏堦抳偟偰偄傞偙偲偑妋偐傔傜傟偨丅傑偨丄傾僆僉偺屄懱慡懱傪悅捈幉傪拞怱偵夞揮偝偣偰杮棃偺岦偒偲堘偆曽岦偵岦偗偨傜偳偆側傞偐傪寁嶼偟偰傒傞偲丄応崌偵傛偭偰偼係侽亾傕庴岝検偑掅壓偡傞偙偲偑暘偐偭偨丅偨偟偐偵丄偦偺応偱偺庴岝検傪戝偒偔偡傞傛偆側懱惂傪偲偭偰偄偨偺偱偁傞丅
梩柺偺妏搙偼丄梩偺宻傊偺晅偗崻偁偨傝偑偹偠傟偰挷愡偝傟偰偄傞偙偲偑懡偄偑丄偐側傜偢偟傕偦偆尷偭偨傢偗偱偼側偄丅偨偲偊偽丄抧忋偺宻傪怢偽偝偢偵抧柺偐傜捈愙梩傪怢偽偡僔僟偺拠娫偱偼丄偦偺梩偼嵶偐偔愗傟崬傫偱暋悢偺晹暘偵傢偐傟偰偄傞暋梩偱偁傞偙偲偑懡偄丅宻偺傛偆偵尒偊傞偺偼梩偺拞墰偺幉丄拞幉偱偁傞丅僔僟偺応崌丄偙偺拞幉偐傜巬暿傟偟偰偄傞彫晹暘傪摿偵塇曅偲傛傇丅椦偺壓偵惗堢偡傞僔僟偱偼丄偙偺塇曅偑扨埵偲側偭偰岝偺曽岦偵庴岝柺傪岦偗偰偄傞乮恾俀乯丅偦偺條巕傪娤嶡偡傞偵偼偟傖偑傫偱墶偐傜尒傞偲傛偄丅塇曅摨巑偑廳側傜側偄偱忋偐傜棃傞岝傪庴偗傞傛偆側丄偪傚偆偳奒抜忬偺峔憿偵側偭偰偄傞偺偑傛偔暘偐傞丅墴偟梩傗恾娪偺奊偩偗偱偼暘偐傜側偄丄岻傒側俁師尦偺岺晇偱偁傞丅

| 恾俀丂 僔僟偺暋梩傪峔惉偡傞塇曅偺妏搙挷愡丅 奺塇曅偑偦傟偧傟偵悈暯偵側偭偰嬻偐傜偺岝傪庴偗偰偄傞條巕偑尒偊傞丅 |
偄偭傐偆丄傎偐偺怉暔偵偍偍傢偢偵柧傞偄偲偙傠偵梩傪峀偘偰偄傞怉暔傪尒傞偲丄偦傟偧傟偺梩偑婲偒忋偑偭偨傝悅傟壓偑偭偨傝偟偰偄偰悈暯偱偼側偄偙偲偑懡偄丅偙傫傕傝偲娵偔栁偭偨怉偊崬傒偺峀梩庽偺奜懁偲撪懁偺巬傪尒斾傋偰傒傞偲椙偔暘偐傞丅
傑傢傝偵岝傪偝偊偓傞傕偺偑側偄偺側傜丄悈暯偵梩傪傂傠偘傟偽堦斣偨偔偝傫岝傪庴偗傜傟傞偼偢側偺偵丄側偤傢偞傢偞偦偭傐傪岦偄偨傝偡傞偺偐偲傕巚偆偑丄梩偼岝偑嫮偔側偭偰偔傞偵偮傟偰岝僄僱儖僊乕傪慡晹偼巊偄愗傟側偔側傞丅傑偨丄恀拫偺懢梲偺捈幩岝偺傛偆偵偁傑傝偵嫮偡偓傞岝傪惓柺偐傜庴偗傞偲丄梩偺壏搙偑忋偑傝偡偓偨傝丄悈偑忲敪偟偡偓偰姡憞偟偰偟傑偭偨傝丄偁傞偄偼夁忚側僄僱儖僊乕傪傕偰偁傑偟偰嵶朎撪偺岝崌惉僔僗僥儉偑偄偨傫偱偟傑偭偨傝丄偲偄偭偨奞傕偍偙傝摼傞丅懢梲岝傪惓柺偐傜庴偗側偄傛偆側妏搙偵梩傪曐偰偽丄嫮岝偺奞傪旔偗傞偙偲偵側傞偟丄傛傝掅偄偲偙傠偵摨偠屄懱偺梩偑偁傞側傜偽丄偦偺梩偲岝傪暘偗崌偭偰岠棪傛偔棙梡偡傞偙偲偵傕側傞丅
娐嫬偵墳偠偰抁帪娫偺偆偪偵梩偺岦偒傪曄偊嫮偄岝傪旔偗傞偲偄偆婍梡側庬椶傕傔偢傜偟偔側偄(3)丅僋僘偼偦偺傛偄椺偩丅僋僘偼丄嬻偒抧傗椦墢偱墵惙偵偮傞傪怢偽偡懡擭惗偺儅儊壢偺偮傞怉暔偱偁傞丅偦偺梩偼俁偮偺彫晹暘乮彫梩乯偐傜側傞暋梩偱偁傞丅偦傟偧傟偺彫梩偼丄栭娫偵偼偳傟傕傎傏悈暯偵傂傠偑偭偰偄傞偺偵丄拫娫偵側傞偲偦傟偧傟偵偹偠傟偰懢梲偵懳偟偰偦偭傐傪岦偔丅偙傟傪傓傝傗傝恓嬥偱悈暯偵屌掕偟偰偟傑偆偲丄擔拞偵梩偵摉偨傞岝偑嫮偔側偭偰梩偺壏搙偑忋偑傝偡偓丄岝崌惉偺懍搙偼偐偊偭偰彫偝偔側偭偰偟傑偆丅
岝偼梩柺偱庴偗傞偺偩偐傜丄梩柺愊偼戝偒偗傟偽戝偒偄傎偳傛偝偦偆偩丅偗傟偳傕丄偁傞尷傜傟偨嬻娫偵傗偭偰偔傞岝偺検偼寛傑偭偰偄傞偐傜丄嫹偄偲偙傠偵傗偨傜偲懡偔偺梩傪媗傔崬傫偱傕庴偗庢傞岝偺検偵偼尷傝偑偁傞丅梩傪嶌傞偺偵傕堐帩偡傞偺偵傕偦傟側傝偵嵽椏傗僄僱儖僊乕偑昁梫偩偐傜丄尒曉傝偺彮側偄梩傪媗傔崬傓偙偲偼丄偐偊偭偰怉暔偵偲偭偰儅僀僫僗偵側傞丅梩摨巑偑擔堿偵側傝偡偓側偄傛偆側揔摉側梩偺検偲暲傋偐偨偑偁傞偼偢偩丅
僔儏乕僩(shoot)偲偄偆怉暔妛梡岅偑偁傞丅宻側偄偟偼巬偲丄偦偙偵偮偄偰偄傞梩偲傪傂偲傑偲傔偵偟偰丄慡懱傪僔儏乕僩偲屇傇丅堦杮偺僔儏乕僩偺拞偱丄宻傪怢偽偝偢偵梩偺枃悢偺傒傪傗偨傜偲憹傗偣偽偦傟傜偺梩偼偍屳偄偵廳側傝崌偆偽偐傝偲側傞丅堦曽丄梩偺偁偄偩偵僗儁乕僗傪偁偗傛偆丄偁傞偄偼怴偟偄嬻娫傊偲揥奐偟偰偄偙偆偲宻偽偐傝偵搳帒偟偰丄娞怱偺梩傪嶌傞嵽椏偑巆傜偢偵宻偽偐傝偺僔儏乕僩偵側偭偰偟傑偭偰偼杮枛揮搢偱偁傞丅揔愗側僶儔儞僗偑偲傟偨丄偄傢偽梩暲傃偺傛偄僔儏乕僩傪嶌傞怉暔偺傒偑恑壔偺拞偱惗偒巆偭偰偙傜傟偨偵堘偄側偄丅
偲偙傠偑丄梩偲宻偺斾棪偑戝偒偔堎側傞僔儏乕僩傪嶌傞庬椶偑悢懡偔抦傜傟偰偄傞丅摿偵栘杮怉暔偱丄宻偑傆偮偆偵怢傃傞僔儏乕僩偲丄宻偼偛偔抁偔丄梩偽偐傝偑栁傞僔儏乕僩偺俀庬椶偑偍側偠屄懱偺側偐偵崿嵼偟偰偄傞椺偑傔偢傜偟偔側偄丅偙偺傛偆側暘壔偑偼偭偒傝偟偰偄傞応崌丄宻偑怢傃傞僔儏乕僩傪挿巬(long shoot)丄悺媗傑傝偺僔儏乕僩傪抁巬(short shoot)偲屇傇丅晛捠丄巬暘偐傟偼挿巬偱偺傒偍偙傞丅
僀僠儑僂丄僇儔儅僣丄僇僣儔側偳偱偼擭偵悢倣倣傕怢傃側偄抁巬偑壗擭傕丄帪偵侾侽擭埲忋傕惗偒懕偗丄梩傪偮偗懕偗傞丅偦傟傎偳嬌抂偱偼側偄偵偟傠丄偁偒傜偐偵悺媗傑傝偱巬暘偐傟傕偟側偄抁巬傪嶌傞椺偼丄僽僫丄僒僋儔丄僐僽僔丄傾僆僴僟側偳偄偔傜傕偁偘傞偙偲偑偱偒傞
屄懱撪偵摨嫃偟偰偄傞丄梩偲宻偺僶儔儞僗偑堘偆巬偼丄栘偺榞慻傒嶌傝偲偄偆婡擻偲岝妉摼偲偄偆婡擻偺栶妱暘扴傪偟偰偄傞偺偩偲峫偊傜傟偰偄傞(4)丅挿巬偼怴偟偄嬻娫傪奐戱偟丄巬暘偐傟傪偟側偑傜庽栘偺戝榞傪嶌偭偰偄偔丅抁巬偼偦偺榞傪懌応偵偟偰岝偺妉摼偵愱擮偡傞丅偲傝偁偊偢偼偦偺応偺岝傪庴偗傛偆偲偄偆偲偒偵丄傗偨傜偲宻傪怢偽偡偺偼帒尮偺柍懯尛偄偱偁傞丅岝妉摼偵摿壔偟偨抁巬偺懚嵼傕丄偠傘偆傇傫棟偵偐側偭偨傕偺偺傛偆偩丅
偝偰丄傕偟榞慻傒嶌傝扴摉僔儏乕僩偲岝妉摼扴摉僔儏乕僩偲偄偆栶妱暘扴偑庽栘偵偲偭偰杮摉偵嬶崌偺偄偄傗傝偐偨側傜偽丄懡彮偲傕宻偺怢挿傪墴偝偊婥枴偵偟偨巬偲偄偆偺偼丄傕偭偲懡偔偺庬椶偱傕尒偮偐傞偐傕偟傟側偄丅偦偆峫偊偰丄昅幰偼偼偭偒傝偟偨抁巬偺暘壔偑抦傜傟偰偄側偄忢椢峀梩庽俉庬傪懳徾偵偟偰丄崱擭怢傃偨巬偺宻偺挿偝偲梩柺愊傪挷傋偰傒偨(5)丅偡傞偲丄挷傋偨偡傋偰偺庬椶偱丄宻偼挿偄偑宻挿偁偨傝偺梩柺愊偑彫偝偄偄傢偽娫墑傃偟偨僔儏乕僩偐傜丄宻偼抁偄偑宻挿偁偨傝偺梩柺愊偑戝偒偄梩柺愊廩幚宆偺僔儏乕僩傑偱偺傂傠偑傝偑偁傞偙偲偑暘偐偭偨丅宻偺挿偝偑俀攞丄俁攞偲挿偄僔儏乕僩偱傕丄梩柺愊偑偙傟偵斾椺偟偰俀攞丄俁攞偲偼側偭偰偄側偄丅偣偄偤偄宻挿偺0.3忔偐傜0.7忔掱搙偵斾椺偡傞偵偲偳傑偭偨丅偪傚偆偳挿巬丄抁巬偲摨條側峔憿偺暘壔偱偁傞丅
榞慻傒嶌傝扴摉僔儏乕僩偲岝妉摼扴摉僔儏乕僩偺懚嵼偑柧傜偐偵側偭偨偲偙傠偱師偵嫽枴偑傕偨傟傞偺偼丄偳傫側忦審偱偳偺傛偆側榞慻傒傪嶌傞偺偐丄偁傞偄偼偁傜偨側榞慻傒傪嶌傜偢偵尰忬偵巭傑偭偰岝傪庴偗傞偺偵愱擮偡傞僔儏乕僩傪嶌傞偺偐丄偲偄偆僟僀僫儈僢僋側惉挿偺夁掱偱偁傞丅巆擮側偑傜偙偺揰偵偮偄偰偼偄傑偩傑偲傑偭偨媍榑偑偑偱偒傞傎偳偵偼尋媶偼恑傫偱偄側偄丅崱屻偺壽戣偱偁傞丅
宻偺拞偱梩摨巑偑偳偺偖傜偄擔堿偵偟崌偆偐偵娭偟偰偼丄梩偺柺愊偲宻偺挿偝偺傎偐偵丄梩偺宍偦偺傕偺傕偢偄傇傫塭嬁偡傞丅抁巬偺傛偆偵宻偑抁偄応崌偵偼丄梩偺宍偵墳偠偰偍傛偦壗枃偖傜偄側傜屳偄偵廳側傝崌傢偢偵峀偘傜傟傞偐偑寛傑傞丅抁巬偱側偔偲傕丄巬愭偵壗枃傕偺梩偑廤拞偟偨僔儏乕僩傪嶌傞怉暔偱偼丄傛傝懡偔偺梩傪偮偗傞傕偺傎偳丄偲側傝偁偆梩摨巑偑廳側傝崌傢側偄傛偆側嵶挿偄宍傪偟偰偄傞孹岦偑偁傞傛偆偩丅傑偨丄堦杮偺宻偐傜弌傞梩偺偁偄偩偱暱乮梩暱乯偺挿偝偵曄壔偑偁傝丄偙偺偨傔偵梩摨巑偺廳側傝崌偄偑旔偗傜傟偰偄傞庬椶傕懡偄丅儎僣僨丄僇僋儗儈僲丄傾僆僊儕丄僷僷僀儎側偳偼丄慡懱偲偟偰嶱偺傛偆側宍偵梩偑攝楍偟偰偄傞丅傑偨丄僀僠儑僂偺抁巬偱傕梩暱偺挿偝偑偨偄傊傫堘偆梩偑崿嵼偡傞乮恾俁乯丅
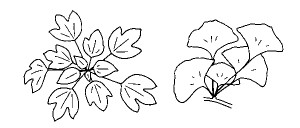
| 恾俁丂 梩暱偺挿偝偑堎側傞偨傔廳側傝崌偄偑旔偗傜傟偰偄傞僇僋儗儈僲乮嵍乯偲僀僠儑僂乮塃乯偺梩丅 |
梩偺宍偺廳梫惈偼梩偑巬愭偵廤拞偡傞応崌偵尷傜側偄丅摨偠柺愊偺梩偑摨偠挿偝偺宻偺忋偵丄摨偠枃悢丄摨偠娫妘偱偮偄偰偄偰傕丄梩偺宍師戞偱梩摨巑偑擔堿偵偟偁偆掱搙偼曄壔偡傞丅梩偺柺偑宻偺嬤偔偵廤拞偡傞傛偆側宍傪偟偰偄傟偽丄宻傪幉偲偡傞嵶偄墌拰偺拞偵梩柺愊偑媗傔崬傑傟傞偙偲偵側傝丄偦傟偧傟偺梩偼擔堿偵側傝偑偪偲側傞丅偄偭傐偆丄偦傟偧傟偺梩偺柺偑宻偐傜墦偄偲偙傠傑偱峀偑偭偰偄傟偽丄傛傝懢偄墌拰偺側偐偵梩柺傪暘嶶偡傞偙偲偵側偭偰丄梩摨巑偑偠傖傑偵側傝偵偔偄丅偦偺偨傔偵偼丄梩傪嵶挿偔偡傞偐丄偁傞偄偼挿偄梩暱傪偮偗傞偺偑桳岠偩乮恾係乯(6)丅屄乆偺梩偺宍偵傕丄宻偺忋偱偺攝楍偲偄偆暥柆偵偍偄偰堄枴偑偁傞崌棟惈偑傒偄偩偝傟傞丅
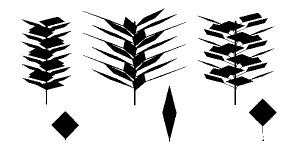
| 恾係丂 僔儏乕僩傪峔惉偡傞梩偺宍偲愯桳嬻娫偺峀偑傝丅 暆峀偱暱乮梩暱乯偺側偄梩偺応崌丄嫹偄嬻娫偵梩偑媗傑偭偰偄傞乮嵍乯丅梩偑嵶挿偄応崌乮拞乯傗梩暱偑偁傞応崌乮塃乯偵偼丄傛傝峀偄嬻娫偵梩偑峀偑傞丅 |
怉暔偺惉挿偼丄岝崌惉婍姱偺帺屓嵞惗嶻偺僾儘僙僗偲偲傜偊傞偙偲偑偱偒傞丅敪夎偟偰嵟弶偵偱偒偨梩偺傑傑偱堦惗傪夁偛偡傛偆側怉暔偼傑偢偄側偄丅嵟弶偺梩偑岝崌惉傪偍偙側偭偰桳婡暔傪壱偓丄偙傟傪巊偭偰偁傜偨偵梩偑嶌傜傟偰屄懱慡懱偺梩偺憤柺愊偼戝偒偔側傞丅梩柺愊偑戝偒偔側傞偲丄傛傝懡偔偺桳婡暔傪壱偓丄偦傟傪尦庤偵偝傜偵愝旛搳帒傪廳偹丄偲偄偆偔傝曉偟偱侾僌儔儉傕側偄庬巕偐傜傕崅偝悢廫儊乕僩儖偺戝栘偑偱偒偁偑傞丅
梩偺柺愊傪憹傗偡偲偄偭偰傕丄宻偺堦屄強偐傜弌傞梩偺悢傪傗偨傜偲憹傗偟偰傕偩傔偱偁傞丅慜偺愡偱傕怗傟偨傛偆偵丄梩摨巑偑擔堿偵偟偁偆偽偐傝偱丄慡懱偲偟偰庴偗庢傞岝偺検偵偼尷傝偼偁傞丅傂偲偮偺夝寛朄偼宻傪偳傫偳傫怢偽偟側偑傜怴偟偄梩傪嶌偭偰偄偔偙偲丄傕偆傂偲偮偼梩傪戝偒偔偡傞偙偲偱偁傞丅堦枃堦枃偺梩傪戝偒偔偟偰丄宻偐傜傛傝墦偔傑偱庴岝柺傪峀偘傟偽丄梩摨巑偑擔堿偵側傞偺傪旔偗側偑傜屄懱慡懱偺庴岝柺傪戝偒偔偡傞偙偲偑偱偒傞丅
偙傟傜偺曽朄偱傕偨偟偐偵奼戝嵞惗嶻偼壜擻偩丅偟偐偟丄懡偔偺庽栘偼巬暘偐傟偵傛偭偰峀偄嬻娫傪僇僶乕偡傞榞慻傒傪嶌傞偲偄偆曽朄傪偲偭偰偄傞丅侾師尦偺懌応偐傜俁師尦偺懌応傊偺僗僥僢僾傾僢僾偩丅忋偵怢傃傞偩偗偱側偔墶偵傕巬傪怢偽偟丄偦偺巬偑偝傜偵暘巬偡傞丅偙傟傜偺巬傪懌応偲偟偰丄偨偔偝傫偺梩傪廳側傝崌偄傪旔偗側偑傜揥奐偡傞偙偲偑偱偒傞丅
儎僔偺拠娫丄僶僫僫偺拠娫丄僜僥僣偺拠娫丄栘惈僔僟側偳偵偼丄挿偝偑悢倣偲偄偆嫄戝側梩傪嶌傞庬椶偑傔偢傜偟偔側偄丅偙傟傜偺怉暔偼丄偄偢傟傕宻偑傎偲傫偳巬暘偐傟偟側偄丅偄偭傐偆偱丄巬暘偐傟峔憿偑敪払偡傞怉暔偱偼偙傫側嫄戝側梩傪偮偔傞傕偺偼偄側偄丅偳偆傗傜丄巬暘偐傟偣偢梩傪戝偒偔偡傞僞僀僾偲丄梩偺戝偒偝偼墴偝偊偰巬暘偐傟偱奼戝偡傞僞僀僾偵暘偗偰峫偊傞偙偲偑偱偒偦偆偩丅
偝偰丄嬻娫偺妋曐偵桳岠側暘巬偩偑丄枅擭擇枖丄擇枖偲暘巬傪孞傝曉偟偰偄偭偨傜偳偆側傞偩傠偆偐丅侾侽擭屻偵偼侾侽侽侽杮偁傑傝丄俀侽擭屻偵偼侾侽侽枩杮埲忋丄俁侽擭屻偵偼侾侽壄杮偺巬偑嶌傜傟傞偙偲偵側傞丅傕偪傠傫丄幚嵺偺怉暔偱偼偦傫側柍拑嬯拑側偙偲偼偍偙傜側偄丅側偤偩傠偆丠
傛偔栁偭偨栘偺庽姤乮梩傪偮偗偨巬偺廤傑傝慡懱乯偺側偐偱栚傪偙傜偟偰傒傞偲丄偁偪偙偪偵屚傟偨巬偑偁傞偺偑暘偐傞丅屚傟偨巬傕偟偽傜偔偼庽忋偵偲偳傑偭偰偟偩偄偵晠偭偰偄偒丄傗偑偰棊偪傞丅戜晽偑捠傝夁偓偨偁偲偵奜傪曕偔偲丄庽忋偵偨傑偭偰偄偨屚傟巬偑傑偲傑偭偰棊偪偰偄傞偺傪尒傞偙偲偑偱偒傞丅
庽忋偺屚傟巬偼丄庽姤偺昞柺偺柧傞偄偲偙傠偵偼彮側偔丄庽姤偺撪晹傗壓偺傎偆偵懡偄丅巬偑崿傫偱埫偔側偭偰偒偨偲偙傠偱偼丄巬偺娫堷偒偑帺慠偵偍偙偭偰偄傞丅傑偨丄埫偄偲偙傠偱偼怴偟偄巬偺敪惗傕峊偊傔偱偁傞丅柧傞偄偲偙傠偺巬偐傜偼壗杮傕丄帪偵偼悢廫杮傕巕巬偑暘巬偟偰偄傞偺偵丄埫偄偲偙傠偺巬偐傜偼傎傫偺侾丄俀杮偟偐弌側偄丄偲偄偆椺偼偄偔傜傕娤嶡偱偒傞乮7)丅
偙偆偟偨巬偺娫堷偒偲嶻帣惂尷偺僾儘僙僗偼丄偄偢傟傕崬傒偡偓傪旔偗傞儊僇僯僘儉偲偟偰摥偔偼偢偱偁傞丅柧傞偄偲偙傠偺巬偑埫偄偲偙傠偺巬傪彆偗傞側偳偲偄偆偙偲傪偣偢丄偁偊偰屄乆偺巬偑棙屓揑偵尒偊傞傛偆側怳傞晳偄傪偡傞偙偲偱丄晄嵦嶼晹暘傪愗傝幪偰丄屄懱慡懱偲偟偰柧傞偄偲偙傠偵梩傪揥奐偡傞偙偲偑偱偒傞丅埫偄偲偙傠偺巬傪傓傝偵堐帩偡傞偨傔偵帒尮傪巊偄丄偦偺偨傔偵傛偄応強偱偺巬偺揥奐偑偍傠偦偐偵側偭偰偟傑偭偰偼屄懱慡懱偵偲偭偰儅僀僫僗側偼偢偩丅
傕偪傠傫丄偙偺傛偆側娐嫬偵埶懚偟偨巇慻傒偩偗偱側偔丄傕偲傕偲堚揱揑偵僾儘僌儔儉偝傟偰偄傞巬暘偐傟僷僞乕儞傕丄偁傜偐偠傔崬傒偡偓傪旔偗傞傛偆側傕偺偱傕偁傠偆丅壗夞偐暘巬傪廳偹偨巬偑丄娐嫬偵偐偐傢傜偢偦傟埲忋偼暘巬偟側偔側傞椺側偳傪偁偘傞偙偲偑偱偒傞丅偦傟偱傕丄廃埻偺岝娐嫬偵廮擃偵懳墳偟偰柧傞偄偲偙傠偵巬傪挘傞偵偼丄嬊強揑側柧埫偵墳偠偨巬偺惗巰傗巬暘偐傟偺挷愡偑桳岠側偼偢偩丅
偲偙傠偱丄庽栘偺巬暘偐傟峔憿傪寁嶼婡偺側偐偱嵞尰偡傞偙偙傠傒偑埲慜偐傜峴傢傟偰偒偨丅杮懡媣晇巵偺巇帠偼偦偺愭嬱偗偱偁傞(8)丅巵偺儌僨儖偱偼丄堦杮偺恊巬偐傜偼俀杮偺巕巬偑弌傞偲偟丄恊巬偐傜巕巬偑弌傞妏搙偲丄恊巬偲巕巬偺挿偝偺斾棪傪僷儔儊乕僞偲偟偰梌偊傞丅偙傟傜傪曄偊傞偙偲偱偝傑偞傑側庽宍偑尰傟偰偔傞丅
昅幰偼丄忋偵弎傋偨巬偺娫堷偒偲嶻帣惂尷偺僾儘僙僗傪杮懡儌僨儖偵慻傒偙傫偱丄寁嶼婡偺拞偱惉挿傪僔儈儏儗乕僩偟偰傒偨(9)丅偦傟偧傟偺巬偵偮偄偰偄傞梩偑偦偺応偺岝娐嫬偵墳偠偰岝崌惉惗嶻傪峴偄丄偦偺寢壥桳婡暔傪偨偔偝傫壱偘偨傜偽偨偔偝傫偺巕巬傪嶌傝丄彮偟偟偐壱偘側偐偭偨傜彮偟偩偗巕巬傪嶌傝丄傕偭偲壱偘側偐偭偨傜巬偛偲屚傜偡偲偄偆夁掱傪孞傝曉偡偺偱偁傞丅偡傞偲丄栘偑惉挿偟偰戝偒偔側偭偰偄偭偰傕巬偺晄帺慠側崬傒崌偄偼偍偙傜側偐偭偨丅傑偨丄庽姤偺側偐偵偼懡偔偺栘偱幚嵺偵娤嶡偝傟傞傛偆側梩偺側偄嬻摯偑偱偒偰偒偨乮恾俆乯丅埫偄偲偙傠偺巬偑屚傟偰偄偭偨寢壥偱偁傞丅

| 恾俆丂 庽栘偺暘巬峔憿偺僔儈儏儗乕僔儑儞丅 巬偛偲偺岝娐嫬傪寁嶼偟偰偦傟偧傟偺巬偺惗巰偲惉挿傪寛傔偨丅庽姤偺抐柺恾乮壓乯偱偼丄奺巬偵偮偄偰偄傞梩傪墌偱帵偡丅庽姤撪晹偵梩偺側偄嬻摯偑偱偒偰偄傞偙偲偑暘偐傞丅 |
庽栘偵偍偗傞晄嵦嶼晹暘偺愗傝幪偰傪暿偺帇揰偐傜尒偰傒傛偆丅怉暔偲摦暔偺戝偒側堘偄偲偟偰丄摦暔偼堏摦偱偒傞偑怉暔偼堏摦偱偒側偄偙偲偑偁偘傜傟傞丅偦傟偼偟偽偟偽丄怉暔偼崻偭偙傪抧柺偐傜堷偒敳偄偰曕偒弌偡偙偲偼偱偒側偄丄偲偄偆宍偱昞尰偝傟傞丅怴偟偄応強傪奐戱偡傞偵偼丄怴偟偄婍姱傪嶌偭偰怢偽偡偟偐側偄丅怴揤抧傊偺堷墇偟偑嬯庤側怉暔偼丄戅媝傕傑偨嬯庤偱偁傞丅摦暔偼偪傚偭偲峴偭偰條巕傪傒偰丄偙傟偼傑偢偄偲敾抐偟偨傜堷偒曉偡偙偲偑偱偒傞丅怉暔偵偼偱偒側偄丅怴偟偄応強偑傛偄応強偐偳偆偐偼偦偙傑偱巬傪怢偽偟偰傒側偄偲暘偐傜側偄丅怢偽偟偰偟傑偭偨巬偼傂偭偙傔傜側偄丅傕偟怢偽偟偨愭偺娐嫬偑岲傑偟偄傕偺偱側偐偭偨傜丄偦偺巬偼偄偝偓傛偔曻婞偟偰偄傞偺偩丅怉暔側傝偺帒尮扵嶕偺傗傝偐偨偱偁傞丅
椬傝崌偭偰惗妶偡傞怉暔偨偪偼丄嫟捠偺帒尮偺庢傝崌偄傪偟偰偄傞丅抧忋偱偼岝傪丄抧壓偱偼悈傗塰梴暘傪傔偖偭偰嫞憟偑偍偙偭偰偄傞丅偁傞屄懱偑帒尮傪庢傞偲偦偺暘傎偐偺屄懱偺庢傝暘偑尭偭偰偟傑偆僛儘僒儉丒僎乕儉偱偁傞丅
怉暔偺嫞憟傪尋媶偡傞傂偲偮偺曽朄偲偟偰丄暋悢偺怉暔屄懱傪堦弿偵堢偰偰傒偰惉挿傪斾妑偡傞偲偄偆傾僾儘乕僠偑偁傞丅堦弿偵堢偰傞偙偲偱挊偟偔惉挿偑埆偔側傞傕偺丄偁傑傝塭嬁偑側偄傕偺側偳偑偄傟偽丄偦傟傪傕偭偰嫞憟偵庛偄丄嫮偄偲偄偆媍榑偑偱偒傞丅捈愙懳寛偝偣偰偦偺寢壥傪尒偰偄傞偺偩偐傜妋偐偩丅偟偐偟丄彑偭偨傕偺偼側偤彑偭偨偺偐丄晧偗偨傕偺偺攕場偼側傫側偺偐偵偮偄偰偺忣曬偼摼傜傟側偄丅嫮偄偐傜彑偭偨丄庛偄偐傜晧偗偨偲偄偆偺偱偼摨媊岅斀暅偱偁傞丅
嫞憟偱偺嫮庛娭學傪傛傝怺偔棟夝偡傞偨傔偵丄梩偺岝崌惉擻椡側偳屄乆偺婍姱偺惗棟揑側惈幙傪挷傋傞偲偄偆傾僾儘乕僠傕偁傞丅偙偺娐嫬忦審偺傕偲偱偼偙偪傜偺傎偆偑偙傟偩偗岝崌惉偺擻椡偑偡偖傟偰偄傞丄偩偐傜偙偪傜偑彑偭偨偺偩丄偲偄偆傛偆偵媍榑偡傞偺偱偁傞丅偨偟偐偵屄乆偺婍姱偺擻椡偼惉挿偺婎杮偱偁傞丅偟偐偟丄偦傟偼偄傢偽僗億乕僣偺抍懱嫞媄偱偺慖庤侾恖侾恖偺塣摦擻椡偺傛偆側傕偺偱丄偦傟偩偗偱偼帋崌偺揥奐傪棟夝偡傞偵偼晄廩暘偩丅
怉暔偺岝傪傔偖傞抧忋晹偺嫞憟偱偼丄嬻娫偺扗偄崌偄偙偦偑彑晧偺杮幙偱偁傞丅傛傝崅偄偲偙傠偺傛傝峀偄嬻娫傪妋曐偡傞偙偲偑彑晧偺寛傔庤偩丅梩偺岝崌惉擻椡偺懡彮偺堘偄傛傝傕丄梩偑忋偵側傞偐壓偵側傞偐偺堘偄偺傎偆偑偼傞偐偵戝偒偔塭嬁偡傞丅堦搙忋偵弌偨屄懱偼朙晉側岝傪棙梡偟偰偳傫偳傫岝崌惉惗嶻傪峴偄丄偝傜偵惉挿偟偰偁傜偨側嬻娫傪妉摼偡傞偙偲偑偱偒傞丅偄偭傐偆丄堦搙擔堿偵側偭偰偟傑偭偨屄懱偼壱偓傕偨偐偑抦傟偰偍傝丄偁傜偨偵宻傪怢偽偡偙偲傕傑傑側傜側偄丅忋傪暍偭偰偄傞偠傖傑側怉暔偑巰偸偺傪懸偮偖傜偄偟偐媡揮偺庤偼側偄丅
怉暔偺抧忋偱偺憡屳嶌梡傪棟夝偡傞偵偼丄偦傟偧傟偺屄懱偑偳偺傛偆側俁師尦峔憿傪偲偭偰偄傞偺偐丄偦偺寢壥丄岝帒尮偺嬻娫暘晍偑偳偺傛偆偵側傞偺偐乮偳偙偑偳傟偩偗埫偔側傞偺偐乯丄偦偟偰丄偦傟偑奺屄懱偺岝偺妉摼偲偝傜側傞俁師尦峔憿偺惉挿偵偳偆塭嬁偟偰偄偔偺偐...偲偄偆僒僀僋儖傪棟夝偟側偔偰偼側傜側偄(10)丅
慜愡偱徯夘偟偨庽栘偺惉挿儌僨儖偼丄屄乆偺巬偛偲偵庴偗庢傞岝偺検傪寁嶼偟丄偦傟偵墳偠偰巬偺惉挿傗惗巰傪寛傔偰偄傞偐傜丄岝偺嬻娫暘晍偲怉暔偺峔憿偲偺僟僀僫儈僢僋側憡屳嶌梡傪嵞尰偱偒傞丅堦屄懱偵尷傜偢丄偨偔偝傫偺屄懱傪偄偭偟傚偵堢偰偰岝偺扗偄崌偄傪偝偣傞偙偲傕偱偒傞丅寁嶼婡偺拞偵椦傪嶌傞偺偱偁傞丅偡傞偲椦偺拞墰晹偺栘偱偼壓偺傎偆偺巬偑屚傟偰偄偔偟丄椦墢偺栘偱偼丄椦偺奜傊岦偐偆巬偼傛偔栁傞偄偭傐偆偱椦偺撪晹偵柺偟偨懁偱偼壓巬偑椦撪摨條偵屚傟偰偄偔乮恾俇乯丅偙偆偟偨宍懺偼丄偄偢傟傕柧傞偄偲偙傠偵巬傪偼偭偨丄屄懱慡懱偲偟偰偮偛偆偺傛偄宍偩偑丄栘慡懱偑柧傞偄曽岦傪嶡抦偟側偔偲傕巬偛偲偺岝娐嫬偵懳墳偟偨巬偛偲偺惉挿斀墳偩偗偱偙偆偟偨庽宍偑偱偒偰偟傑偆丅庽姤撪偺晄嵦嶼晹暘偺愗傝幪偰偼丄屄懱偺偁偄偩偱岝傪扗偄崌偭偰偄傞偲偒偵丄柧傞偄娐嫬傊巬偲梩傪廤拞偝偣傞儊僇僯僘儉偲偟偰摥偔丅

| 恾俇丂 俁俇杮偺儌僨儖庽栘傪奿巕忬偵攝抲偟偨応崌偺僔儈儏儗乕僔儑儞丅 扨撈偱堢偰傞偲墌悕宆偵惉挿偡傞儌僨儖庽栘傕丄椦偺拞偱堢偰傞偲乮嵍乯丄拞墰偺屄懱偱偼壓巬偑屚傟偁偑傝乮拞乯椦偺墢偵抲偐傟偨屄懱偱偼椦偺奜偺柧傞偄懁偺巬偺傒偑敪払偡傞乮塃乯丅乮TAKENAKA 1994b傛傝夵曄乯 |
恾俇偺傛偆側庽宍偼幚嵺偺栘偱偛偔偛偔晛捠偵娤嶡偱偒傞丅椦偺側偐偺崅栘偱偼壓偺埫偄偲偙傠偺巬傪棊偲偟偰偄傞偟丄椦偑摴楬傗嬻偒抧丄愳側偳偵柺偟偰偄傞偲偙傠偱偼丄柧傞偄懁偺巬偑敪払偟偰偄傞丅
偁傞峔憿偑偳傫側栶偵棫偭偰偄傞偐丄偳傫側婡擻傪壥偨偟偰偄傞偐偽偐傝傪尒偰偄偰傕丄怉暔偺惗偒曽偺棟夝偼敿偽偵偲偳傑傞丅側偵傪偡傞偵傕僞僟偲偄偆偙偲偼側偔丄側傫傜偐偺僐僗僩偑偲傕側偆丅偁傞偄偼丄偙偪傜傪棫偰傟偽偁偪傜偑棫偨偢丄偲偄偆僕儗儞儅偑偁傞丅怉暔偼宍偱彑晧偡傞偲偄偭偰傕傗傝偨偄曻戣傪偱偒傞偺偱偼側偄丅偝傑偞傑側惂尷偺側偐偱僶儔儞僗傪偲傝側偑傜偺彑晧偱偁傞丅
偨偲偊偽丄懡彮偲傕抧柺偐傜忋偵梩傪帩偪忋偘傞偵偼丄暔棟揑側巟偊偲偟偰傕悈偺嫙媼側偳惗棟揑側巟偊偲偟偰傕宻偑昁梫偩(11)丅宻偼丄悈摴娗偺傛偆側暔幙桝憲娗偺栶妱偲揝嬝偺傛偆側巟拰偺栶妱偲傪寭偹偨僷僀僾偵偨偲偊傞偙偲偑偱偒傞丅偙偺宻傪偳偺偖傜偄偺懢偝偵偡傞偐丄偲偄偆偺偼怉暔偑忢偵捈柺偡傞栤戣偩丅懢偔偡傟偽偡傞偩偗埨慡偱偼偁傞偟丄悈傕廩暘偵嫙媼偱偒傞丅偟偐偟丄偦傟偩偗嶌傞偨傔偺嵽椏傗堐帩偺僐僗僩傕戝偒偔側傞丅傑偨丄偁傞懢偝偵偡傟偽愨懳埨慡偲偄偆傕偺偱偼側偄丅傑傟偵悂偔嫮偄晽傗丄忋偐傜棊偪偰偒偨懠偺怉暔偺屚傟巬丄摜傒偮偗偰偄偔摦暔側偳丄偳傫側徴寕偑偄偮棃傞偐暘偐傜側偄丅傑偨丄宻傪挿偔怢偽偣偽廃埻偺屄懱傛傝忋偵弌傜傟傞偐傕偟傟側偄偑丄庤帩偪偺桳婡暔偵偼尷傝偑偁傞偲偒偼丄怢偽偣偽怢偽偡傎偳懢偝傪媇惖偵偟偰埨慡棪偑掅偔側傞丅偳偙傜傊傫偱愜傝崌偄傪偮偗偨傜偄偄偺偐丄怉暔偼寛抐傪敆傜傟傞丅
峔憿傪嶌傞偆偊偱偳偺傛偆側僕儗儞儅偑偁傞偺偐丄偦偺拞偱偳偺傛偆偵僶儔儞僗傪偲偭偰惗偒偰偄傞偺偐丄側偳傪掕検揑偵柧傜偐偵偟偰偙偦怉暔偑偳偺傛偆偵寛抐偟丄偳偺傛偆偵彑晧偟偰偄傞偺偐傪傎傫偲偆偵棟夝偟偨偙偲偵側傞偼偢偩丅怉暔偺惗妶傪棟夝偡傞偨傔偵傕丄偝傜偵偼孮棊傗怷椦偺側偐偱偺怉暔摨巑偺憡屳嶌梡傪棟夝偡傞偨傔偵傕丄峔憿偲婡擻偲傪摑崌偟偰峫偊偰偄偔昁梫偑偁傞偩傠偆丅偙傟偐傜偺尋媶壽戣偼懡偄丅
掕検揑側尋媶偼傑偩偠傘偆傇傫偵偼恑傫偱偄側偄偑丄怉暔偺庬椶偵傛偭偰偝傑偞傑側惗妶偺巇曽偑偁傞偺偲懳墳偟偰丄愜傝崌偄偺偮偗偐偨偵傕偝傑偞傑側僷僞乕儞偑偁傞傛偆偩丅偨偲偊偽丄崱擭偟偐惗偒傜傟側偄堦擭憪偼彨棃傪婜懸偟偨惗偒曽傪偡傞偼偢偼側偔丄懠偺怉暔偵暍傢傟偨傜偽埨慡棪偼屻夞偟偵偟偰傔偄偭傁偄攚怢傃傪偡傞丅偄偭傐偆丄嫞憟偵彑偮偙偲偼偁偒傜傔偰擔堿偵側偭偰傕偟傇偲偔曢傜偡惗偒曽偵揙偡傞椦彴偺掅栘側偳偱偼僐儞僷僋僩偱僐僗僩偑偐偐傜側偄宍傪嶌傞丅傑偨丄擔堿偵側偭偰傕徟傜偢偵偠偭偲変枬偟偮偮嵟廔揑偵偼崅栘偵側偭偰斏怋偡傞偲偄偆惗偒曽傪偡傞栘偱偼丄埨慡丒妋幚巜岦偺惉挿僷僞乕儞傪帵偡丅摿偵庽栘偱偼丄慜偵弎傋偨榞慻傒嶌傝扴摉僔儏乕僩偲岝妉摼扴摉庬僔儏乕僩偺嶌傝傢偗偲攝抲偺偟偐偨傕偙偆偟偨愜傝崌偄偺偮偗偐偨偺廳梫側億僀儞僩偵堘偄側偄丅
怉暔偺偐偨偪偺側偐偵偳傫側崌棟惈偑傂偦傫偱偄傞偺偐丄偄偐偵怉暔偼偐偨偪偱彑晧偟偰偄傞偺偐傪弎傋偰偒偨丅偲傝偁偘偨榖戣偼丄偩傟偱傕帺暘偺栚偱妋偐傔傞偙偲偑偱偒傞傛偆側偙偲偽偐傝偱偁傞丅偙偺傎偐偵傕丄怉暔偼偝傑偞傑側崌棟惈傪暥帤捠傝"懱尰"偟偰偄傞丅怉暔偼摦暔偲堘偭偰摝偘偨傝偟側偄偐傜丄偄偔傜偱傕婥偺偡傓傑偱挱傔傞偙偲偑偱偒傞丅偤傂丄偦偺尗偝傪尒偮偗偰傗傠偆偲偄偆栚偱尒夞偟偰丄妝偟傫偱偄偨偩偗偨傜偲巚偆丅怉暔偨偪偑偦傟偧傟偵僕儗儞儅傪夝寛偟偰偄傞偄偆偙偲偵傑偱巚偄傪傔偖傜偣傟偽丄側偍偺偙偲嫽枴怺偄偼偢偱偁傞丅