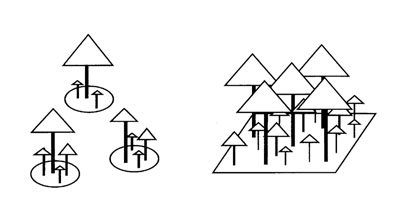
| 図1 森林のギャップモデル(左)と3次元連続空間モデル(右).ギャップモデルは一本の成木が占める面積を単位とする.それらの間の相互作用は考えない場合が多い. |
竹中明夫(国立環境研究所・生物圏環境部)
Updated on 14 March 2000
空高くから見下ろせば緑の平面としか見えない森には,たくさんの木々が生活している.森だけ見て木を見なければ,森の中で起こっているさまざまな営みも,その総体としての森林のダイナミクスも十分には理解できない.一方,一本の木を見つめるばかりで,その木が森の一員としてまわりの木々や動物,物理環境と相互作用をしていることを忘れてしまったら,その木の生活の理解も不十分なものとなろう.木を見て森も見ることで,両者の理解が深まるはずだ.
複雑なシステムを研究対象とするとき,ひたすら要素へと還元する道を進むばかりでは,システム全体の理解へはたどり着かない.要素についての理解にもとづきながら,システムをモデル的に再構成することの重要性についてはいまさら述べるまでもない.本講演では,森林というシステムを相互作用しあう木々の集まりとしての表現する,個体ベースモデルと呼ばれるモデルについて述べる.個体ベースモデルとはどのようなものであり,また,森林がどのように個体ベースモデルで表現されてきたかを概観する.ついで,私が現在開発中の,個体ごとの遺伝情報を組み込んだモデルを2つ紹介する.
個体ベースモデルは,多くの生物個体からなるシステムを表現するモデルの一種である.生物は動物であっても植物であってもよいし,単一種からなるシステムでも多種のシステムでも表現できる.一個体一個体を識別して取り扱うことがその特徴である.集団が個体のあつまりであることを意識したモデルであっても,個体数,サイズ分布,年齢分布などで集団を表現するものは個体ベースモデルとは呼ばない.個体1はこんな性質の持ち主,個体2はあんな性質,という情報を保持しながらそれぞれの個体の挙動を扱うのが個体ベースモデルである.全体の個体数やバイオマスだけを扱うモデルに比べると,個体ベースモデルは多量の計算を必要とする場合が多い.当然コンピュータの利用が不可欠である.近年,個体ベースモデルを利用した研究がしだいに広がりつつある背景には,コンピュータの高速化と普及がある.
計算の手間をかけても個体ベースのモデルを使う意義はどこにあるのだろうか.システムの登場人物がそれぞれに個性を持っていたり、登場人物間の相互作用がおたがいの位置関係に依存するような場合には、全体を平均してしまうとシステムの本質が見えなくなってしまうことがある.また,個体数の有限性が本質的な意味を持つようなシステムも,集団を連続変数であらわすモデルでは表現しにくい.個体を識別しないモデルでは表現が難しいプロセスが,システム全体の挙動と本質的にかかわっているような場合には,個体ベースモデルが活躍する.
森林は個性ある登場人物が集まり,相互の位置関係に依存した相互作用をおこなっているシステムである.種類が違う木は成長速度も死亡のしかたも違う.個体によって高さが違い,高い木は低い木を日陰げにする.低い木にとっては,近くの高木は光を遮る邪魔者だが遠くの高木の影響は受けない.種子は親の木の近くに多く散布される.遠くの木よりは近くの木と多くの花粉をやりとりする.さらに,種の多様性に関心があるならば局所的な絶滅過程を再現したいが,そのためには個体数の有限性を無視するわけにはいかない.こうして並べてみると,まさに個体ベースモデルは森林を表現するのにうってつけのモデルのようである.
もちろん,個体を認識しない森林のモデルがだめなモデルだということではない.たとえば,大気と生物圏との水,熱,二酸化炭素のやりとりを表現するモデルでは,植物群落全体が均一な一枚の葉であるかのような単純化をすることが多い.巨視的に見たときにほぼ均一な植物の層が覆っているような場合には,これでじゅうぶん目的にかなったモデルであり,個体性を持ち込んで不必要に複雑化することはない.
一般に,モデルは複雑すぎても困るし単純化しすぎても困る.複雑になればなるほど計算の手間もかかるし個々のプロセスや要素とシステム全体とのつながりが見えにくくなってしまう難点もある.いっぽう,単純化しすぎて,現実世界で直接観察できる「こと」や「もの」との対応のつけようがなくなってしまったモデルもまた利用しにくい.システムのどのような面を再現したいのかを踏まえつつ,適切な単純化をする必要がある.
森林の個体ベースモデルの代表的なものは,ギャップモデルと呼ばれる一群のグループである.このモデルは,十分に成長して林冠(森林上部の枝葉が茂っている部分)に到達した木一本が占める面積をひとつの単位としている.その面積の中での個体間の日陰にする・されるの関係に基づいてそれぞれの木が受ける光の量を計算し,木の成長速度を決めてゆく.木の成長や死亡の特性は,樹種ごとのパラメータとして与えられる.単位となる区画が林冠ギャップ(一本の高木が死んでできる林冠の空隙)に対応するものであることから,ギャップモデルと呼ばれる.ギャップができたあとその下の若木が成長してギャップを埋める林冠木の交替のプロセスは、ギャップ更新と呼ばれる.個体の上下関係に基づいて受光量を決め,それが成長に反映されるモデルでは、ギャップ更新がおのずと表現される.高木が死ねばその下が明るくなり,それまで日陰にいた小個体の成長が促進されてギャップを埋めていくのである.
ギャップモデルでは数十平方メートル程度を単位面積とすることが多い.光の奪い合いを計算するときには,ひとつの単位面積内での木の高低の関係のみが考慮される.すなわち単位面積内では1次元モデルとなっている.単位区画の集合として森林を表現するが,単位区画の位置関係や相互作用は考慮しないモデルが多い(図1左).
ギャップモデルの最初のものは,Botkinら(1972)が開発したJABOWAモデルである.それについで,Shugart(1977)の FORETも発表された.それ以来,世界各地の森林を対象に,数多くのモデルが作られてきた.それらの基本構造は,みなJABOWAモデルやFORETモデルを踏襲している.ギャップモデルの歴史についてはHuston (1992),Shugart and Smith (1992), Liu and Ashton (1995) などが詳しい.
|
| 図1 森林のギャップモデル(左)と3次元連続空間モデル(右).ギャップモデルは一本の成木が占める面積を単位とする.それらの間の相互作用は考えない場合が多い. |
多くのギャップモデルは森林の平面的な広がりを考慮していないが,近年は3次元の連続空間をそのまま扱うモデルも作られるようになった(図1右).そのなかではPacala et al.(1993)によるSORTIEが有名である. Pacalaらは、北米の落葉広葉樹と針葉樹からなる森林を対象に、そのおもな構成樹種9種の成長と死亡の特性を3年間にわたって調査した.その結果をもとに開発された個体ベースモデルSORTIEは3次元連続空間モデルであり,個々の木の光環境は近隣の木による被陰を逐一評価して決めている.
組み立てたモデルは,現実のシステムをうまく再現するかどうか検証する必要がある.再現できなかったらその理由を検討しなくてはならない.要素の組み立て方がまずいのかもしれないし,要素についての知識が正しくないのかもしれない.あるいは重要な要素を見落としているのかもしれない.モデルとしての組み立て方のまずさは純粋にモデルの世界の問題となるが,要素についての知識の正しさや本質的な要素を見落としていないかどうかに関しては,モデルそのものにとどまらず,これまで研究者がそのシステムをどのようにとらえ定量化してきたかという部分も問われることになる.
PacalaらのSORTIEは,実際の森林で観察される種組成の時間変化をよく再現しており,SORTIEの基本構造と,樹種ごとの特性を表すパラメータが,おおむね妥当なものであることを示唆している.いっぽう,SORTIEの内部で空間的な光環境の不均一性を平均化してしまうと,種組成の時間変化が正しく再現できなくなり,森林のバイオマスの見積もりも現実の値から大きくはずれてしまった.このことは光の空間的不均一性の重要性を示すものである. 個々の木が近隣の木に日陰にされる効果を計算することで,SORTIEは現実の森林の動態をよく再現し得ている.
一般に,森林では個体間の光をめぐる競争がきわめて本質的な要素であり,多くの個体ベースモデルがなんらかの形でこのプロセスを組み込んでいる.明示的に光の奪い合いを扱わないモデルであっても,ある面積に生存可能な林冠木を1本と限るような形で,光資源による個体数の制限を組み込んでいる.
個体ベースモデルを実用的なツールとして利用していくためには,メンテナンスの容易さと扱いやすさが重要な要素となる.目的に応じた改変が容易であることは,実用ツールとして利用していく上では必須の条件である.
オブジェクト指向のプログラム設計は,近年広く行われつつある技法である.プログラムをデータ処理のアルゴリズムを中心に設計するのではなく,そのプログラムが扱う'こと'や'もの'の表現方法を中心に設計しようというものである.現実世界の'こと'や'もの'をコンピュータのなかに再現しようというシミュレーションのプログラミングとは非常に相性がよさそうである.個体という'もの',すなわちオブジェクトの振る舞いを記述する個体ベースモデルではなおさらである.システムの構造を素直に表現するように設計されたオブジェクト指向のプログラムは,あとからの改変や拡張がやりやすい.
生態学の数値計算シミュレーションモデルの世界でも,しだいにオブジェクト指向の設計が広まりつつある.現在はまだ,オブジェクト指向の手法を使うこと自体が研究のひとつのセールスポイントになるような段階だが,遠からずしてごくあたりまえの技術となるだろう.
現在,わたしは2つの個体ベース森林モデルを開発している.ひとつは3次元の連続空間モデル,もうひとつは空間を格子に区切って一区画には一個体しか生育させないという単純な格子モデルである.
まず,連続空間モデルGEFLIの概略を紹介する(図2).GEFLIの登場人物は,それぞれいくつかのデータのまとまりで表現される.たとえば木は樹種、サイズ、森林内の位置、光環境などのデータで表現される.SORTIEと同様に,木の相互の位置関係にもとづいてそれぞれの木が受ける光の量を決める.木の上からあちこちを見上げてみて,空が見えるならばその方向からの光が届き,視線が他の木に遮られるならばその方向からの光は届かない.木の成長,死亡,種子の生産は、樹種、サイズ、光環境などに依存して決める.その依存性の詳細は樹種ごとにパラメータとして与える.
このモデルには送粉者、種子捕食者,そして伐採,山火事,病害などの撹乱要因も組み込むことができる.土壌条件などの空間的な不均一性を組み込むことも可能だ.もちろん、必要に応じて組み込み得るということであって,あまり重要ではないと判断される場合には無視すればよい.要素の足したり引いたりが柔軟にできるところは,高度に抽象化せずに現実をなぞるモデルの利点である.
木々の挙動を詳細に再現しようとすると,与える必要のあるパラメータの数はどうしても多くなる.すべてのパラメータを測定に基づいて決めていくのはたいへんな時間と労力がかかるし,現状では測定がきわめて困難なパラメータもある.とはいえ,各パラメータとシステムの挙動との関係を調べることでさまざまな情報が得られるだろう.
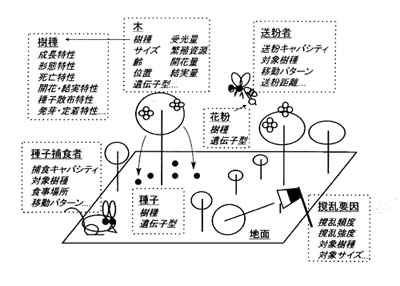
| 図2 森林の個体ベースモデルGEFLIに組み込まれた要素(竹中1999より引用) |
GEFLIがこれまでのモデルと異なるのは,それぞれの木が遺伝子を持ち,これが親から子へと遺伝するプロセスが組み込まれている点である.開花した木が作る種子には,その種子をつくった親木の遺伝子と,花粉を供給した個体の遺伝子とが半分づつ含まれている.
集団遺伝学は生物集団の中での遺伝子の頻度の変化を研究対象とする.これまでの集団遺伝学は,集団を構成する個々の生き物がどのように空間に配置されているかを考慮してこなかった.しかし、動き回ることができない植物の集団内での遺伝子のやりとりは,個々の植物の位置関係に強く影響されるだろう.近年、DNAの塩基配列に基づく親子判定が可能になるなど,植物集団の遺伝的構造について得られる情報は飛躍的に多くなってきている.これまで接点が少なかった集団遺伝学と生態学との境界領域は,学問的にも大きな発展の可能性を持つ.さらに,生物の絶滅過程での遺伝的な多様性の消失,遺伝的な劣化などの重要性が注目されつつある現在,生物多様性の保全のためにも重要な分野となる. GEFLIの狙いのひとつは,この境界分野に切り込むことである.
GEFLIを使ったシミュレーションの例として、種の局所的な絶滅と林の面積,そして繁殖様式の関係を調べた結果を紹介する.25種類の木からなる森林を考えた.これらの樹種の特性はまったく同じに設定した.外部からの種子の供給はないので,仮想森林からいったん姿を消した樹種は復活せず,局所的な絶滅が起こる.共存を促進するような特段のメカニズムは組み込まれておらず,確率的に絶滅が発生し,時間とともに残存種数は減少していく.成木の寿命が100年前後になるようなパラメータ設定のもとで,800年間にわたって木々の成長をシミュレートしたところ,絶滅の速度は森林の面積に強く依存することが確かめられた(図3左).
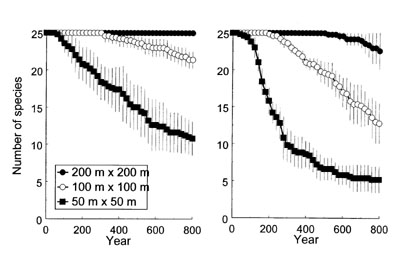
| 図3 個体ベース森林モデルGEFLIでシミュレートした,25種の樹木からなる仮想森林での絶滅による種数の減少. 森林の面積は3通りに設定し、それぞれ5回の試行を行った.死亡確率●、○、■は各面積での試行の平均、縦線は標準偏差を示す.(左)自家和合性の場合,(右)自家不和合性の場合.(竹中1999より引用) |
次に、25種の樹木がそれぞれ自家不和合性の性質を持つ場合を考えた.自家不和合性は、めしべの側と花粉親の側の遺伝的な同一性によって種子の稔実率が低下する現象である.近親交配を避けるための仕組みとして機能するが,集団の個体数が減少して自家不和合性を支配する遺伝子の多様度が低下すると,めしべ側と同じ遺伝子型の花粉が来る確率が高まって種子ができにくくなり,個体数の減少にいっそう拍車がかかる可能性がある.自家不和合性を支配する遺伝子が8種類ある場合を想定してシミュレーションをおこなってみたところ、不和合性がない場合とくらべて絶滅速度が明らかに大きくなった(図3右).
上に紹介した連続空間モデルはたいへんリアルであり,特定の目的に特化していないだけに拡張も容易である.いっぽうで,さまざまなプロセスを詰め込みすぎると,パラメータの数の多さ,ある要素とシステム全体の動きとの関係の見通しの悪さ,計算時間がかかるなどの難点も生じる.モデルを利用して調べたいことがらがはっきり絞られているならば,その目的に特化したモデルを利用するほうが得策である.
私が開発中のもうひとつのモデルLatFor-Gは,成長や死亡の詳細な再現はせず,種多様性と遺伝的多様性のダイナミクスのシミュレータに特化している.個体間の光の奪い合いのプロセスを再現するには,大量の計算をしなくてはならない. LatFor-Gでは光をめぐる競争を直接扱わないため劇的に計算量が減り,必要な演算時間はGEFLIに比べて2ケタから3ケタも短くなる.地面は二次元の格子に区切られ,各マス目には一本の木しか生育しない.あるマス内の個体が死亡したら,近くの個体が散布する種子から新しい個体を発生させて空いたマスを埋める.花粉を提供する個体は,種子を供給する親個体の近くの開花個体から選ぶ.新個体が両親の遺伝子型を半分づつ受け継ぐことは先のモデルGEFLIと同様である.樹種の特性を表すパラメータは,種子の散布パターン,花粉の散布パターン,成熟までの年数と最大寿命,死亡率,そして自家不和合性の程度など少数に絞られる.
林の形を自由に作れることと各個体が遺伝情報を持つことを生かしたシミュレーション実験を行ってみた.有限の個体数しかいない集団では,確率的に種の絶滅や遺伝子の消失が起こる.林の形に応じて種の局所的な絶滅や遺伝子の消失の速度がどう変化するかを調べた.大きなひとかたまりの林がある場合,これが8つの小さな林に分割された場合,そして分割された小さな林のあいだが廊下のような幅の狭い木立ちでつながれている場合を想定した(図4).この仮想森林には2種類の木が共存している.木の最大寿命は100年で,樹齢20年から種子をつけ始める.種子は親木から最大4マス,花粉は5マス離れたところにまで散布される.この散布距離では,8つに分割された林の間のすきまを飛び越えることはできない.

| 図4 格子上の個体ベース森林モデルLatFor-Gによるシミュレーション実験に用いた森林の空間配置.40×20マスの大きな仮想森林(左),10×10マスの小さな林8つに分けた場合(中央),小さな林の間を6×2マスの廊下状の木立でつないだ場合(右). 2種類の木を混在させてある.樹冠の濃淡の違いは種類の違いを表す. |
分断された森林を回廊でつなぐと,ひとつの林で局所的な絶滅や遺伝的な多様性の消失が起こっても,隣接する他の林から種子や花粉が供給されやすく,多様性を保つうえで効果があるのではないかと考えられている.5000年にわたるシミュレーション結果はこの予測とよく一致するものだった.すなわち,(1)小さい林では高い確率で2種のうち1種が絶滅した.しかし,小さい林を廊下でつなぐことで絶滅の確率は大きく低下した(図5左).(2)大きな林に比べて,小さな林の中での対立遺伝子の数(遺伝子の種類数)は急速に減少した.しかし,いくつもの小さい林を廊下でつなぐことで,それぞれの林の中での遺伝子の減少速度は大きく低下した(図5右)
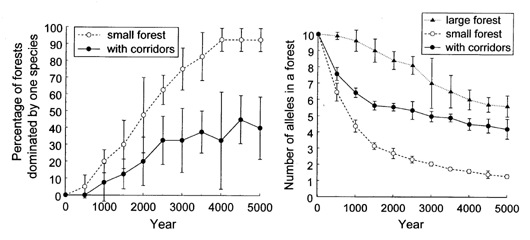
| 図5 格子上の個体ベース森林モデルLatFor-Gによるシミュレーション実験の結果.(左)2種の樹木が共存していた林のうち1種が絶滅したものの割合の時間変化.(右)林の中の個体群中に維持されている対立遺伝子(allele)の種類数の時間変化.それぞれ5回の試行の平均値と標準偏差を示した. |
この結果は当然と言えば当然であり,あえてシミュレーション計算をしなくとも定性的には想像がつく.しかし定量的な予測が求められる場面では,あることに「効果がある」というだけでは不十分であり,「どれだけ効果があるか」を提示しなくてはならない.具体的な数字の答えを出すためには,ここで紹介したようなシミュレーションモデルが活用できる.保護区を孤立されるよりも回廊でつないだほうがよいだろうということは想像がつくが,どのぐらいよいのか,また,回廊の太さや長さに依存してその効果はどのように変化するのか,そうした具体的な数値を出せるという点で,このようなモデルには実用的な価値がある.
モデルは決して万能ではない.個別の知識を並べただけでは得られない情報を引き出せるとは言っても,個別の知識自体が不完全なものである以上,それらを統合したモデルもおのずと不完全なものである.また,自然が本来持っている確率性ゆえにモデルの予測も確率的であらざるをえない.それでも,個体ベースモデルはこれからますますその活躍の場を広げていくと思われる.学問の世界のみならず,環境アセスメントや自然保護地域の設定などの場面での利用も多くなるだろう.ただ,ひとくちに個体ベースモデルと言ってもその構造はさまざまである.リアルにしようとすればするほど複雑になるが,いたずらに複雑にしてもいけない.かといってあまりに単純化しすぎてもまずい.モデルを使って表現したいこと,再現したいことは何なのか,そのためにはどのような要素が必須なのかを見極めて,適切なモデルを設計しなくてはいけない.また,構成要素からシステムを再構成するモデルは,構成要素の振る舞いについての知識を充分に生かすものでなければならない.森林の個体ベースモデルにおいても,木々の生活についての研究の蓄積を生かしてこそ,木の理解と森の理解をつなげるものとなるだろう.
Botkin, D.B., Janak, J.F. & Wallis, J.R. 1972. Some ecological consequences of a computer model of forest growth. J. Ecol. 60:849-872
Huston, M. 1992. Individual-based forest succession models and the theory of plant competition. in "Individual-based models and approaches in ecology" (eds. DeAgnelis, D.L. Gross, L.J.) Chapman & Hall, New York, London. pp408-420.
Liu, J. and Ashton, P.S. 1995. TL:Individual-based simulation models for forest succession and management. For. Ecol. Manage. 73:157-175.
Pacala, S.W., Canham, C.D. and Silander, Jr., J.A. 1993. Forest models defined by field measurements: I. The design of a northeastern forest simulator. Can. J. For. Res. 23:1980-1988.
Pacala, S.W. and Deutchman, D.H. 1995. TL:Details that matter: the spatial distribution of individual trees maintains forest ecosystem function. Oikos 74:357-365.
Shugart, H.H.and West, D.C. 1977. Development of an Appalachian deciduous forest succession model and its application to assessment of the inpact of the chestnut blight. J. Environ. Manage. 5:161-179..
Shugart, H.H. and Smith, T.M. 1992. The potential for application of individual-based simulation models for assessing the effects of global change. Annu. Rev. Ecol. Syst. 23:15-38:
竹中明夫 1999. 木々の知識を集めてみれば:個体ベースモデルによる森林の動態の再現と予測.保全生態学研究 4:135-141.