以下は、 保全生態学研究会 発行の「保全生態学研究」Vol.4(2) に掲載の私の文章を、同研究会事務局の了解を得て公開するものです。
森林の将来を予測するとき、基礎データを集めたあとはエキスパートが長年の経験とカンで判断しますというのでは、科学的な議論の土俵には乗らない。仮定と計算プロセスが明らかにされたモデルによる予測なら、その当否を科学の言葉で具体的に論じることができる。そのようなモデルを積極的に利用することで環境アセスメントはより科学的なものとなるはずだ。本稿では森林を表現するモデルのひとつである個体ベースモデルを紹介し、科学的な環境アセスメントの道具としての利用価値について考える。
森林は多様な登場人物が複雑に相互作用するシステムである。いろいろな種類の植物はもちろんのこと、そこに暮らす動物も、土壌環境や微気象も、相互作用の複雑な網の目に絡まりあっている。ちょっと眺めたぐらいでその動きや将来が読めてしまうような単純なシステムではない。
そのような対象に対し、生態学者はなんとかシステムの本質をつかみ取ろうとしてきた。まずは構造を記載するところから始める。どんな種類のどんな大きさの木が、どのように配置されているのかを調べる。そこにはなんらかのパターンがあるかもしれない。
といっても、森林の構造はけっして静的なものではない。つねに誕生、成長、繁殖、死亡といったプロセスが起こっているダイナミックなシステムだ。生態学者は、このシステムのありかたと密接にかかわっていると睨んだ要素に注目して研究してきた。複雑な全体を理解するために、部分に分解して調べてみようというアプローチである。
プロセスを解析したらそれで森林が分かってしまうかというと、そうはいかない。個々の知識をもういちど統合して、森林というシステムを再構成してみることが必要である。個々のプロセスを定量的に調べても、そのあとの議論がすべて定性的なものにとどまるならば、現実の系と照らし合わせることも、また応用の道も限られたものになる。プロセスの知識を統合して定量的な数値計算モデルを作り、システム全体がそれらしく再現されるならば、本質的な要素を押さえたのだと考える根拠が得られる。
再現されたモデル森林は、生態学者の見た森林のエッセンスをかたちにしたものである。また、そのようなモデルはさまざまな仮想実験に利用することができる。その応用の道は多い。環境アセスメントのための利用はその筆頭にあげることができよう。
森林でおこっているプロセスを統合する枠組みとして適当と思われるのが個体ベースモデルである。このモデルは、森林全体で木が何本だとかバイオマスが何トンだとかいったひとまとまりにした扱いをするのではなく、一本一本の木が成長し、死亡し、繁殖する様子をコンピューターの中に再現するものだ。ばらばらに並べただけでは何を語っているのかよく分からない木々の知識を集めて、森林というシステムを再構成する。
森林は個性ある登場人物が相互作用をしているシステムだ。個々の木は異なる樹種であったり、異なるサイズであったり、あるいは異なる遺伝子型であったり、とそれぞれに個性がある。光などの資源をめぐる競争は、位置関係とサイズの大小に依存する相互作用である。種子は親木のまわりに散布され、近くの同種個体とのあいだでは花粉を介した遺伝子の交流がある。
登場人物がそれぞれに個性を持っていたり、登場人物間の相互作用がおたがいの位置関係に依存したりするようなシステムでは、個体ベースモデルが有効なことが多い。そのようなシステムでは、全体を平均してしまうとシステムの本質が見えなくなってしまうことがままある。森林全部をひとまとめにするモデルに比べるとはるかに複雑になる個体ベースモデルだが、これなしでは表現しきれない側面を持つのが森林である。
最初の森林の個体ベースモデルは1972年に発表されたJABOWAモデルである(Botkin, Janak & Wallis 1972)。このモデルは、森林を一本の高木の樹冠がおおう面積程度の小区画に分けてその中での被陰関係のみを考え、小区画のあいだの相互作用は考慮していない。その後、これに類するモデルはいくつも作られた(Shugart & Smith, 1992) 。最近では、森林全体の3次元構造をそのまま表現して個体間の相互作用を取り扱うモデルも作られるようになった(Pacala et al. 1996)。
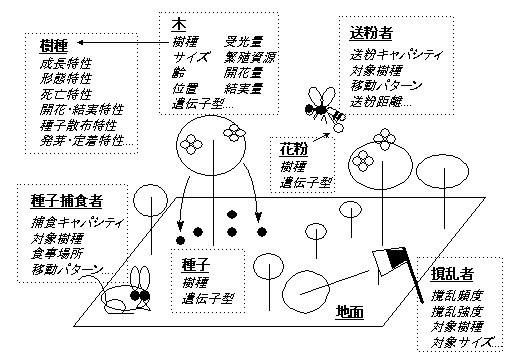
現在わたしが開発中のモデルGEFLIの登場人物たちを図1に示す。各登場人物は、この図に示したようなデータのまとまりとして扱われる。たとえば個々の木は樹種、サイズ、森林内の位置、光環境などのデータをあわせて表現される。
図1 個体ベースモデルGEFLIに組み込んだ要素の模式図
光環境は個々の木のサイズと位置関係にもとづいて計算する。一本一本の木の上に視点を置いて、そこから空のあちこちを見上げる。その視線が他の木に遮られたら、その方向からの光は来ないはずだ。小さい木の近くに大きな木が居るならば、小さい木には光はとどきにくくなり、その木は暗い環境で暮らすことになる。
木の成長(サイズの増加)、種子の生産、死亡などのプロセスは、樹種、サイズ、光環境、そして必要なら遺伝子型に依存させる。明るいほどよく成長し、死ににくく、たくさん種子を作るのが一般的だが、その依存性の詳細は、樹種やサイズや遺伝子型に応じて決める。とくに光環境に注目するのは、となりあう木々が樹冠を接するような森林では、光をめぐる競争が個体間の相互作用としてもっとも重要だからである。
木々のあいだの競争関係は、A種とB種ではA種が強い、という形でその帰趨を決めるのではない。個々の木は光環境に依存して挙動を決めるし、また、それぞれの木の存在は、他の木をとりまく光環境に影響する。個体間の競争はすべて光環境を介して記述される。
高木が死んで林冠に空隙(ギャップ)ができたあとその下の若木がぐんぐんと成長してギャップを埋める、という高木の交替のプロセスは、ギャップ更新と呼ばれる。個体のサイズと位置に基づいて日陰にする、されるの関係を決めていくモデルでは、この現象もおのずと表現される。高木が死ねば、その近くにいた小さい個体の光環境は改善する。明るくなれば成長が早まり、ギャップを埋めていくのである。
このモデルには撹乱要因、送粉者、種子捕食者も組み込める。撹乱要因は人による伐採でもいいし、山火事や台風、崩落、あるいは病害なども表現できる。空間構造を持つモデルだから、土壌条件の不均一性を組み込むことも可能だ。もちろん、組み込みたければ組み込めるとこうことであり、無理に全部を考慮する必要はない。これらが森林のありかたに密接に関与している場合には組み込めばよいし、あまり重要ではないと判断すれば無視すればよい。関係する要因を足したり引いたり柔軟にいじれるところは、高度に抽象化せずに現実をなぞるモデルの利点である。
空間構造を持つモデルでは、森林の面積や形も明示的に扱われる。保護区のサイズや形態を変えて実験することもできる。土地は不連続であってもよい。小さな森林を細い回廊でつなぐことの効果も調べられるだろう。取り組むべき課題にあわせてさまざまな実験が考えられる。
花が咲いて受粉すると種子ができる。花粉は送粉者が運ぶ。送粉者は風であってもよい。両親の遺伝子型の組み合わせによって種子ができにくくなる不和合性も組み込める。種子は両親の遺伝子型を引き継ぐ。散布された種子から新しい個体が発生する。一定頻度で空から種子が降ってくるタイプのモデルと異なり、親木の数が減少すれば散布される種子の数は減り、親木がゼロになれば種子の供給もゼロになる。絶滅プロセスを再現するにはこのようなモデルが適当だ。
このモデルの大きな特徴は、各個体に遺伝子型をあらわすデータを持たせたことだ。個体の振る舞いが遺伝子型によって変わるようにすれば、自然選択による遺伝子頻度の変化をシミュレートできる。遺伝子が個体の振る舞いにまったく影響しないとすれば、偶然による遺伝子頻度の変化、すなわち遺伝的浮動を観察することもできる。
これまでの集団遺伝学では、集団の空間構造をほとんど考慮してこなかった。しかし、固着性の生物である植物の集団の遺伝的な構造を考えるとき、空間構造が重要な意味を持つであろうころは想像にかたくない。近年、DNAのマイクロサテライト領域を調べることで、集団の遺伝的構造について飛躍的に多くの情報を得られるようになってきた。その成果を森林で起こっていることの理解につなげていくうえでも、空間構造を持った個体ベースモデルは有力な道具だ。
遺伝子をモデルに組み込むことは、進化生態学や集団遺伝学の興味に応えるいっぽうで、保全生態学の観点からも大きな意味がある。遺伝的多様性が失われる過程、自家不和合性や近交弱性の影響、外部から持ち込まれた個体による遺伝的汚染など、遺伝学の観点から指摘されている保全上の問題点について考える道具となるはずだ。
個体ベースモデルのメリットのひとつとしてあげられることは、森林で直接目に見えるプロセスを無理に単純化せずに取り込んでいるために自然のなかで調べられた木々の知識をそのまま生かしやすい点である。Pacalaら(1996)は、北米の落葉広葉樹と針葉樹からなる森林を対象に、その主要構成種9種の成長と死亡にかかわるパラメータを3年にわたって測定した。その結果をもとに、空間構造をもった森林の個体ベースモデルSORTIEを開発した。SORTIEは、実際の森林の遷移にともなう種組成の変化をよく再現した。このようにモデルの挙動と現実の森林の動態とを照らし合わせることで、モデルの構造の正当性とパラメータの妥当性を検証をすることができる。
温帯林であれば10種程度ですむものの、1キロ四方に数百種 から1000種もの樹木が生 育している熱帯林など、どうしたらよいだろうか。Koehler & Huth (1998) は、熱帯の400以上の樹種をいくつかの機能的に似通ったグループに分類して、グループごとにひとまとめにして共通のパラメータを与えた。そのモデルが描く森林の動態は、多数の種からなる実際の森林とよく対応するものだった。グループにまとめて扱ってもよいのなら、多様性が高い森林でも対応できる。
コンピュータの中に仮想の森林をつくることのメリットのひとつは、簡単に実験ができることである。開発にともなって森林の面積を半分にすることの影響が論点になっているとき、実際に木を切ってしまっては事前アセスメントにならない。コンピュータの中の森林なら面積を半分にするのも2倍にするのも容易だ。択伐の影響や外来種の導入の影響などもシミュレーションにより予想してみることができる。
実験の例として、わたしが開発したモデルでの絶滅過程のシミュレーションを紹介する。まず、25種の木からなる森林を考える。成長や死亡については、Pacalaら(1996)のパラメータのセットのうち一種のものをそっくり借り、繁殖にかかわる部分については適当にパラメータを与えて森林の種構成の変化を追ってみた。まったく同じ性質の25種の樹木が同数いる状態から出発する。外部からの種子の供給は考えないので、一度絶滅した種はそれっきりであり、確率的に種数は減少していく。特に共存を促進するようなメカニズムは何もない。この確率的絶滅の速度は、当然予想されるように森林の面積に強く依存することが確かめられた(図2)。面積が大きいほど絶滅速度は小さくなり、200m四方の森林では800年たってもまったく絶滅は起こらなかった。

図2 25種の樹木からなる森林での、確率的な絶滅による種数の減少。
個体ベースモデルGEFLIによるシミュレーションの結果を示す。。森林の面積は3通りに設定し、それぞれ5回の試行を行った。死亡確率は樹種、サイズ、光環境により一意的に決めているが、特定個体の生死は乱数により決めるため、試行ごとに結果は違う。●、○、■は各面積での試行の平均、縦線は標準偏差を示す。
次に、自家不和合性を組み込んでみた。自家不和合性は、めしべの側と花粉親の側の遺伝的な同一性によって稔実率が低下する現象である。植物がそのような性質を持つ場合、個体数が少なくなると遺伝的な多様度が低下し、種子ができにくくなることで個体数の減少が加速され得る。一遺伝子座で8種類の遺伝子がある場合を想定してシミュレーションをおこなってみたところ、予想通り、不和合性がない場合とくらべて絶滅速度がはっきりと大きくなった(図3)。
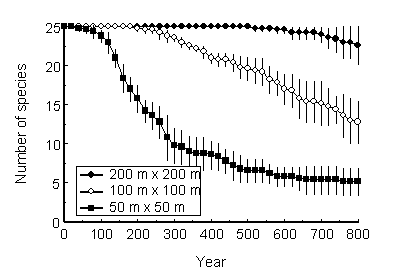
図3 図2の設定に、自家不和合性を組み込んだ場合の種数の減少過程。
シミュレーションに必要なパラメータをすべて野外で測定することはかなりの労力を伴う。しかし、パラメータが揃う前の段階であっても、パラメータを少しずつ変化させながらモデル森林の挙動を見ることで、各パラメータの重要性を調べることができる。また、一部にしろ得られたデータを組み込んでその意味を考える助けともなる。そして、データの集積が進めば、より現実的な予測が可能になっていく。
車を運転するとき、一時停止の標識があればいったん止まる。無視したら確実に事故が起こるとは限らないが、事故が起こる可能性はたしかにあるからだ。環境アセスメントの場面でシミュレーションモデルを使う場合にも、同じような心構えが必要だ。自然現象にはつねに偶然の要素があるし、木々の性質を調べるにも誤差は避けられないから、人間が持っている科学的知識をフルに動員して構築した森林モデルといえども、その予測は確率的なものとならざるを得ない。ある種が絶滅する可能性が50%という予測に対して、確実に絶滅すると証明されたのでないから無視してよいと考えるのは適切ではない。望ましくない事態が起こる可能性があるならば、その可能性の大きさや、どのぐらい望ましくないことなのかを総合して判断する必要がある。そうした心構えを持ち、天気予報のような科学的確率予報の道具として利用するとき、森林のモデルは環境アセスメントに不可欠なものとなると考える。
Botkin, D.B., Janak, J.F. & Wallis, J.R. (1972) Some ecological consequences of a computer model of forest growth. J. Ecol., 60:849-872
Koehler, P. & Huth, A. (1998) The effects of tree species grouping in tropical rainforest modelling: Simulations with the individual-based model FORMIND. Ecol. Model. 109:301-321.
Pacala, S.W., Canham, C.D. & Saponara, J. (1996) Forest models defined by field measurements: estimation, error analysis and dynamics. Ecol. Monogr. 66:1-43.
Shugart, H.H. & Smith, T.M. (1992) The potential for application of individual-based simulation models for assessing the effects of global change. Annu. Rev. Ecol. Syst. 23-15-38: