| 恾侾丂 屄梩偺岝崌惉懍搙偺岝埶懚惈丅丂攋慄偼嫮岝慾奞偑偍偒傞応崌丅 |
怉暔偺偐傜偩偼丄彫偼暘巕偺峔憿偐傜戝偼崅栘偺暘巬峔憿傑偱丄偝傑偞傑側僗働乕儖偺峔憿偑奒憌揑偵慻傒崌傢偝偭偨僔僗僥儉偱偁傞丅偦偟偰丄偳偺僗働乕儖偺峔憿傕怉暔偑惗偒偰偄偔偨傔偵昁梫側婡擻偲側傫傜偐偺偐偐傢傝傪帩偭偰偄傞丅
怉暔偺惗偒曽偺偟偔傒傪棟夝偟傛偆偲偄偆壢妛幰偺搘椡偼丄偟偩偄偵儈僋儘偺僗働乕儖偺峔憿偲婡擻偺夝柧傊偲恑傫偱偒偨丅偦偺惉壥偼戝偒偄丅偟偐偟丄偦偙偱偁偒傜偐偵偝傟偨偙偲偑怉暔偺惗偒曽偺側偐偱帩偮堄媊傪掕検揑偵栤偄側偍偡搘椡偼偐側傜偢偟傕偠傘偆傇傫偱偼側偐偭偨丅怉暔偺棟夝偺偨傔偵偼偠傑偭偨尋媶偑丄慻怐傊丄嵶朎傊丄偦偟偰暘巕傊偲恑傫偩偒傝怉暔傊栠偭偰偙側偄偺偱偼偄偗側偄丅
怉暔偺偐傜偩偺側偐偺偝傑偞傑側僾儘僙僗偺抦幆傪丄傑傞偛偲偺怉暔偲偄偆暥柆偺側偐偵傕偆偄偪偳抲偒捈偡偵偼偳偆偟偨傜傛偄偩傠偆偐丅屄乆偺抦幆傪慻傒崌傢偣偰壖憐揑側儌僨儖怉暔傪嶌偭偰傒傞偺偑桳岠側傾僾儘乕僠偺傂偲偮偩偲巚偆丅偦偺偨傔偵偼寁嶼婡偺棙梡偑晄壜寚偩傠偆丅偦偆偟偰慻傒忋偘偨儌僨儖怉暔偼丄怉暔偺惗偒曽偺巇慻傒偵偮偄偰偺尰帪揰偱偺抦幆傪廤戝惉偟偨傕偺偱偁傞丅偦偺傛偆側儌僨儖傪帩偮偙偲偱丄屄乆偺僾儘僙僗偺慡懱偺側偐偱偺堄媊偯偗傪昡壙偡傞偙偲傕偱偒傞丅
傑傞偛偲偺怉暔傪峫偊捈偡偲偒丄屄乆偺婍姱偺側偐偱偍偙偭偰偄傞偙偲偺扨弮側懌偟嶼偱偼偡傑側偄丅懡偔偺婍姱偑桳婡揑偵慻傒偁傢偝偭偰丄怉暔傪偲傝傑偔娐嫬偺側偐偱偳偺傛偆偵婡擻偟偰偄傞偺偐傪峫偊偹偽側傜側偄丅偦偺傛偆側帇揰偐傜怉暔傪懆偊傞偲偒丄屄懱慡懱偲偟偰偺岝丄悈丄塰梴墫椶側偳偺帒尮偺妉摼僾儘僙僗偼晄壜寚偺梫慺偱偁傞丅
怉暔偑妉摼偟偨帒尮偑懱偺側偐偱棙梡偝傟傞僾儘僙僗偵偮偄偰偼丄偍偍偔偺尋媶幰偑偦偺夝柧偵庢傝慻傫偱偒偨丅偦傟偵偔傜傋傞偲帒尮傪妉摼偡傞傑偱偺僾儘僙僗傊偺拲栚搙偼偄傑傂偲偮偩丅岝娐嫬傊偺墳摎丒弴壔偵偟偰傕丄梩偺側偐偺儗儀儖偱偺尋媶偼悢懡偄偺偵斾妑偟偰丄梩偺拞偵擖偭偰偔傞岝偺検偲怺偔偐偐傢傞抧忋晹偺儅僋儘側峔憿傊偺娭怱偼偠傘偆傇傫偱偼側偐偭偨丅
偟偐偟丄偦傟偱偄偄偺偩傠偆偐丅堦枃偺梩偑丄傎偐偺梩偺擔堿偵側傞偐側傜側偄偐丄偁傞偄偼懢梲偺岝偑傑偭偡偖偁偨傞偺偐幬傔偵偁偨傞偺偐丅偦傫側偙偲偱丄梩偑庴偗偲傞岝偺検偼壗攞傕曄壔偟摼傞丅偦傟傪柍帇偟偰偄偄偼偢偼側偄丅梩偺拞偺岝崌惉僾儘僙僗偺岠棪偑偨偄偣偮側偺偲摨條偵丄屄懱偑岝傪庴偗庢傞岠棪傕偨偄偣偮偩丅
杮峞偺僥乕儅偼丄怉暔偺抧忋晹偺儅僋儘側峔憿偲丄岝傪妉摼偡傞婡擻偲偺娭學偩丅怉暔偺偐傜偩偺屄乆偺僾儘僙僗傪屄懱偲偄偆暥柆偺側偐偱峫偊偰傒傞偆偊偱丄懡彮偲傕嶲峫偵側傟偽岾偄偱偁傞丅
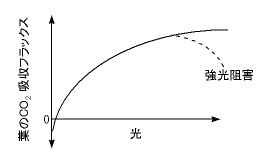
杮戣偱偁傞峔憿偲岝妉摼婡擻偺娭學偵擖傞慜偵丄傑偢丄怉暔偵偲偭偰偼偳偺傛偆側岝偺庴偗曽偑朷傑偟偄偺偐傪峫偊偰傒傛偆丅婎杮偵側傞偺偼梩偺岝崌惉懍搙偺岝埶懚惈丄偄傢備傞岝亅岝崌惉嬋慄偱偁傞乮恾侾乯丅
|
| 恾侾丂 屄梩偺岝崌惉懍搙偺岝埶懚惈丅丂攋慄偼嫮岝慾奞偑偍偒傞応崌丅 |
偦偺摿挜偺戞侾揰偼丄岝崌惉懍搙偑岝偺検偲偲傕偵憹壛偡傞偙偲偱偁傞丅岝偙偦偑僄僱儖僊乕尮側偺偩偐傜摉慠偩丅
摿挜偺戞俀揰偼丄忋偵撌側朞榓宆偺嬋慄偱偁傞偙偲偩丅擖幩岝偁偨傝偺岝崌惉懍搙偼岝偺検偑憹偊傞偲偲傕偵掅壓偟偰偟傑偆丅偝傜偵丄懡偡偓傞岝偼奞偵偡傜側傞丅揹巕揱払宯儗儀儖偱偺晄搒崌傕偍偙傞偟丄擬晧壸偵傛傞梩偺壏搙偺忋徃傗悈暘偺懝幐傕儅僀僫僗偵摥偔偙偲偑偁傞丅偦偺寢壥丄嫮岝偺傕偲偱偐偊偭偰岝崌惉懍搙偑掅壓偟偰偟傑偆嫮岝慾奞傕偍偙傝摼傞丅
堦枃偺梩偵夁忚側嫮岝偑偁偨傞偙偲傪旔偗傜傟傟偽丄怉暔偵偲偭偰俀偮偺儊儕僢僩偑偁傞丅傂偲偮偼丄嫮岝慾奞偵傛傞岝崌惉偺掅壓傪偝偗傞偙偲偱偁傝丄傕偆傂偲偮偼丄岝傪傗傝偡偛偟偰傎偐偺梩偵岝傪傑傢偡偙偲偱屄懱慡懱偲偟偰偺岝崌惉検傪戝偒偔偱偒傞偙偲偱偁傞丅梩偑擖幩岝偵懳偟偰側側傔偵側傞側偳偟偰擖幩岝偑尭傟偽丄偦偺梩偵媧廂偝傟側偄岝偺検偑憹偊傞丅偦偺暘偺岝傪傎偐偺梩偑媧廂偡傟偽岝傪暘偗偁偭偨偙偲偵側傞丅岝崌惉偺岝埶懚惈偑朞榓宆偱偁傞偐傜丄堦枃偺梩偑巊偄愗傟側偄岝傪庴偗偰傕偆堦枃偺梩偑岝晄懌偺忬懺偵偍偐傟傞傛傝傕丄俀枃偺梩偲傕偵暯嬒揑側岝傪庴偗偰偄傞傎偆偑崌寁偺岝崌惉検偼戝偒偔側傞丅摨偠屄懱偺梩偱岝傪暘偗偁偊偽丄屄懱慡懱偺岝崌惉惗嶻偑憹壛偡傞偙偲偵側傞丅
怉暔孮棊偺岝崌惉惗嶻偺棟榑揑側儌僨儖偺尋媶偱偼丄孮棊傊偺擖幩岝偑戝偒偄応崌偵丄梩傪悈暯傛傝傕孹偗偰岝媧廂偺岠棪傪壓偘偨傎偆偑孮棊慡懱偺岝崌惉惗嶻偑戝偒偔側傞偙偲偑帵偝傟偰偄傞(1) 丅偙偺尰徾偺杮幙偼丄傑偝偵岝偺暘偗偁偄偵傛傞岝棙梡岠棪偺夵慞偱偁傞丅孮棊偺忋晹偺梩偑丄岠棪揑偵偼棙梡偟偒傟側偄傎偳偺岝傪撈愯偟偰偟傑偆傛傝傕丄孮棊偺壓偺傎偆傑偱懡偔偺岝傪摟夁偝偣偰岝偺張棟擻椡偵梋椡偑偁傞梩偵暘偗傞傎偆偑慡懱偲偟偰偺惗嶻偑戝偒偔側傞丅
偨偩偟丄傗偨傜偲偨偔偝傫偺梩傪偮偗偰岝傪暘攝偡傟偽傛偄偐偲偄偆偲丄偦偆偱偼側偄丅堦枃偺梩偁偨傝偺岝偑彮側偔側傝偡偓偰偼崲傞丅傎偳傎偳偺枃悢偺梩偱暘偗傞偺偑傛偄丅偳傟偖傜偄偑傎偳傎偳偐偼屄懱偺傑傢傝偺岝娐嫬偵埶懚偡傞丅岝偑傆傫偩傫偵偁傞偲偙傠偺怉暔偱偼懡偔偺梩傪偮偗偰岝傪暘扴偟偰棙梡偡傞偺偑傛偄偟丄椦彴側偳埫偄娐嫬偺怉暔偱偼廳側傝偁偄傪旔偗偰彫悢偺梩傪偮偗丄偦傟偧傟偺梩偑庴偗傞岝偺検傪側傞傋偔戝偒偔偡傞偺偑傛偄偼偢偩丅
屄乆偺梩偺庴岝検傪戝偒偔偡傞偵傕丄懡偔偺梩偺偁偄偩偱側傞傋偔嬒摍偵岝傪暘偗傞偵傕丄怉暔懱偺抧忋晹偺峔憿偑廳梫側栶妱傪偵側偆丅偮偓偺愡偱偼丄怉暔偺儅僋儘側俁師尦峔憿偺掕検揑側懆偊偐偨偵偮偄偰弎傋傞丅
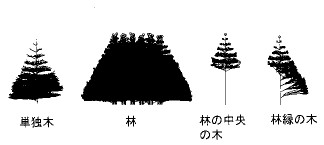
杮峞偱儅僋儘側峔憿偲屇傇偺偼丄擏娽偱擣幆偱偒偰丄庤偵庢偭偰尒傞偙偲偑偱偒丄暔嵎偟傪偁偰偰挿偝偑偼偐傟傞傛偆側僒僀僘僗働乕儖偺峔憿偺偙偲偱偁傞丅側傫偺摴嬶傕側偟偵栚偱尒偊傞偺偩偐傜丄應掕傕偐傫偨傫偺傛偆側婥傕偡傞偺偩偑丄偨偔偝傫偺宻傗梩偑擖傝慻傫偩怉暔偺3師尦峔憿偺應掕偼偦偆梕堈偱偼側偄丅怉暔偺偐傜偩慡懱傪傂偲偐偨傑傝偺暔懱偲偟偰應掕偡傞偺偼崲擄側偺偱丄峔惉梫慺偵暘偗偰偦傟偧傟偺宍偲埵抲傪應掕偡傞偺偑摼嶔偱偁傞丅
怉暔偺梩偺宍偼庬椶偵傛偭偰偍偍傓偹寛傑偭偰偄傞丅揟宆揑側梩偺宍傪墴偝偊偨偆偊偱丄屄乆偺梩偺挿偝偲暆側偳偺僒僀僘僷儔儊乕僞傪應掕偟丄偦傟偵偁傢偣偰揟宆揑側宍傪怢弅偝偣傞偙偲偱屄乆偺梩傪昞尰偱偒傞丅宻偼傕偭偲娙扨偩丅偨偄偰偄偺宻偼墌拰偺慻傒崌傢偣偲傒側偣傞丅墌拰側傜偽挿偝偲懢偝傪應傟偽傛偄丅
偮偓偵丄偙傟傜偺晹昳偺埵抲傪寛傔側偗傟偽側傜側偄丅偦偺偨傔偵偼丄傑偢丄崪慻傒偵側傞宻偵偮偄偰丄抧柺偺偒傢偐傜弌敪偟偰丄傂偲偮偺墌拰偲傒側偣傞嬫夋偛偲偵偦偺椉抂揰娫偺挿偝偲妏搙乮揤捀妏偲曽埵妏乯傪應掕偟偰偄偔丅暔嵎偟丄暘搙婍丄悈弨婍丄僐儞僷僗側偳偺摴嬶偑偁傟偽傛偄丅梩偑偮偄偰偄傞偲偙傠偱偼丄丄宻忋偱偺梩偺埵抲丄戝偒偝丄婎晹偐傜愭抂傊偺揤捀妏偲曽埵妏側偳傪應掕偡傞丅偙偆偟偰摼傜傟偨忣曬傪傑偲傔傟偽丄屄懱慡懱偺嬻娫峔憿偑悢抣壔偱偒偨偙偲偵側傝丄寁嶼婡偺拞偵棫懱峔憿傪嵞峔惉偡傞偙偲傕娙扨偵偱偒傞(2), (3) 丅
嬻娫撪偱偺埵抲傪傛傝娙曋偵應掕偡傞偨傔偺憰抲傕岺晇偝傟偰偄傞丅偨偲偊偽丄昅幰傜偑峫埬偟偨憰抲偼嬥懏惢偺暘搙婍傆偨偮偲姫偒広偲偐傜側偭偰偄傞(4)丅 應掕偟偨偄揰偵傑偱姫偒広傪堷偭挘傞偲丄應掕婍偐傜偺嫍棧偑撉傔傞傎偐丄傆偨偮偺暘搙婍偦傟偧傟偱揤捀妏偲曽埵妏偑撉傒庢傟傞傛偆偵側偭偰偍傝丄憰抲偺拞怱傪尨揰偲偡傞嬌嵗昗宯偺拞偱偺埵抲傪抦傞偙偲偑偱偒傞丅恾俀偼偙偺憰抲傪巊偭偰應掕偟偨僨乕僞偵傕偲偯偄偰昤偄偨忢椢峀梩庽僔儘僟儌偺抰庽偺奊偱偁傞丅
| 恾俀丂 宻偲梩偺俁師尦嬻娫拞偺埵抲忣曬傪巊偭偰昤偄偨僔儘僟儌偺抰庽丅丂 摨偠屄懱傪忋偐傜尒偨偲偙傠乮塃忋乯偲丄懁曽乮俀曽岦乯偐傜尒偨偲偙傠乮壓乯 |
偙偆偟偨偄傢偽儘乕僥僋宯偺應掕曽朄偵懳偟丄僴僀僥僋宯偺曽朄傕偁傞丅偦偺傂偲偮偼帴応傪棙梡偡傞傕偺偱偁傞丅帴応敪惗婍偲丄帴応傪僙儞僗偡傞偨傔偺僾儘乕僽偲偐傜側傞偙偺僔僗僥儉偼丄傕偲傕偲偼寁嶼婡傪巊偭偨僶乕僠儍儖儕傾儕僥傿偱偺巊梡傪憐掕偟偰奐敪偝傟偨傕偺偺傛偆偩丅恖娫偺3師尦嬻娫偱偺摦偒傪寁嶼婡傊偺擖椡偲偟偰悢抣壔丒揹巕忣曬壔偡傞偨傔偵巊偆偺偱偁傞丅偙偺憰抲偱丄帴応敪惗婍偐傜悢倣偺斖埻撪偺應掕偑壜擻偱偁傞丅埵抲傪應掕偟偨偄揰偵僾儘乕僽傪抲偄偰僗僀僢僠傪墴偡偲丄帴応敪惗婍偵懳偡傞憡懳揑側埵抲忣曬偑摼傜傟傞丅屄乆偺梩偺埵抲傗丄宻偺墌拰偲傒側偣傞晹暘晹暘偺椉抂偺埵抲傪應掕偟偰偄偗偽丄怉暔偺俁師尦峔憿偺忣曬偑摼傜傟傞丅
偁傞揰岝尮偺傕偲偵堦屄懱偺怉暔偑偄傞偲偟傛偆丅偦偺怉暔偺堦枃堦枃偺梩偺昞柺偱偺岝僼儔僢僋僗偼丄傆偨偮偺梫慺偵埶懚偡傞丅傂偲偮偼偦偺梩傊偺岝偺擖幩妏丄傕偆傂偲偮偼傎偐偺梩傗宻偵偠傖傑偝傟偰埫偔側傞憡屳旐堿偺桳柍偱偁傞丅
梩偑堦枃偟偐側偄怉暔傗偮傞偱抧柺傪偼偆怉暔偺傛偆偵梩偺憡屳偺斴堿偑側偄応崌偵偼丄梩偑庴偗傞岝偺検偺寁嶼偼梕堈偱偁傞丅岝偲梩柺偲偺妏搙乮兤乯偑暘偐傟偽丄梩柺忋偱偺岝偺擖幩僼儔僢僋僗偼岝偵悅捈側柺偱偺僼儔僢僋僗偵cos兤傪偐偗偨傕偺偵側傞丅岝偲悅捈側梩偑傕偭偲傕柧傞偔徠傜偝傟丄岝偲暯峴側梩偺柺偼傑偭偨偔岝傪庴偗傜傟側偄丅偠偭偝偄偺嬻偺傕偲偱偼丄懢梲偐傜偺捈幩岝偺傎偐嬻偺偁偪偙偪偐傜嶶棎岝偑棃偰偄傞偑丄偙傟傕揰岝尮偺婑偣廤傔偲峫偊偰丄屄乆偺揰岝尮偐傜偺岝偺偁偨傝偐偨傪挷傋偰偐傜愊嶼偡傟偽傛偄丅
暋悢偺梩傪帩偮怉暔懱偺庴岝婡擻傪昡壙偡傞偵偼憡屳旐堿傕峫椂偟側偔偰偼偄偗側偄丅梩堦枃偺応崌偲偔傜傋傞偲偩偄傇柺搢偵側傞偑丄崅峑儗儀儖偺夝愅婔壗偱寁嶼偡傞偙偲偑偱偒傞丅
怉暔偺俁師尦峔憿偵傕偲偯偔庴岝検偺寁嶼曽朄偼丄戝偒偔暘偗偰傆偨偮偁傞丅偙傟傜傪尒壓傠偟朄偲尒忋偘朄偲屇傇偙偲偵偟傛偆乮恾俁乯丅
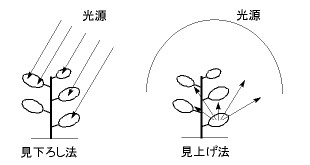
| 恾俁丂 怉暔偺庴岝検傪寁嶼偡傞俀偮偺曽朄丏丂 尒壓傠偟曽乮嵍乯偱偼嬻偺岝尮偐傜岝慄傪偍傠偟偰梩偲傇偮偐傞偐偳偆偐傪寁嶼偡傞丅尒忋偘朄乮塃乯偱偼梩偺忋偵帇揰傪抲偄偰嬻偺偁偪偙偪偵偁傞岝尮傪尒忋偘丄傎偐偺梩偑偠傖傑偵側傞偐偳偆偐傪寁嶼偡傞丅 |
尒壓傠偟朄偱偼丄嬻偐傜怉暔傪尒壓傠偟偰丄偳偺梩偑偳偺偖傜偄尒偊偰偄傞偐傪寁嶼偡傞(3)丅嬻偐傜尒偊偰偄傞晹暘偼丄偡側傢偪嬻偐傜偺岝偑摉偨偭偰偄傞晹暘偱偁傞丅傎偐偺梩偵塀傟偰傒偊側偄晹暘偼岝傪庴偗傜傟側偄丅梩偺尒偊偐偨偼丄摉慠尒傞曽岦偵傛偭偰堘偆丅懢梲偐傜偺捈幩岝偺傒傪峫偊傞側傜丄懢梲偺曽岦偐傜尒壓傠偣偽傛偄偟丄慡揤偐傜偺嶶棎岝傪寁嶼偡傞側傜丄嬻偺偁偪偙偪偐傜尒壓傠偟偨寢壥傪丄嬻偺奺曽岦偺柧傞偝偵偁傢偣偰廳傒偯偗偟偰偐傜崌寁偡傟偽傛偄丅
傕偆傂偲偮偺曽朄丄尒忋偘朄偼丄怉暔偺傎偆偵帇揰傪抲偔曽朄偱偁傞(4)丅梩偺忋偺堦揰偐傜嬻傪尒忋偘偰丄帇慄偑懠偺梩傗宻偵幷傜傟傞側傜丄偦偺曽岦偐傜偺岝偼偦偺帇揰偵偼撏偐偢丄傕偟嬻偑尒偊偰偄傞側傜丄岝偑撏偔偙偲偵側傞丅尒壓傠偟朄偺応崌偲摨條丄懢梲偐傜偺捈幩岝偺傒傪峫偊傞側傜丄懢梲偺曽岦偩偗傪尒忋偘傟偽傛偄偟丄慡揤偐傜偺嶶棎岝傪寁嶼偡傞側傜丄嬻偺偁偪偙偪傪尒忋偘傞昁梫偑偁傞丅
尒忋偘朄偵偟傠丄尒壓傠偟朄偵偟傠丄梩偑庴偗傞岝偺愨懳検傪寁嶼偡傞偵偼丄嬻偺偳偺曽岦偐傜偳偺偖傜偄偺岝偑偔傞偺偐丄偲偄偆岝尮偵偮偄偰偺忣曬偑昁梫偱偁傞丅岝尮偼懢梲偐傜偺捈幩岝偲慡揤偐傜偺嶶棎岝偲偵暘偗偰埖偆偙偲偑懡偄丅
抧媴忋偺偁傞抧揰偐傜尒偨偲偒偺懢梲偺埵抲偼丄婫愡偲帪崗丄偦偟偰偦偺抧揰偺堒搙偵埶懚偟偰寛傑傞丅懢梲偺捈宎偼帇嵎妏搙偵偟偰栺0.5搙偱偁傝丄傎傏揰岝尮偲偟偰埖偆偙偲偑偱偒傞丅傕偆堦曽偺嶶棎岝傕傕偲偼懢梲偐傜偺捈幩岝偱丄偙傟偑戝婥拞偱嶶棎偟偨傕偺偺堦晹偑抧柺偵撏偄偰傞丅嶶棎岝偺暘晍偵偮偄偰偼丄嬻慡懱偵傢偨偭偰嬒堦偵柧傞偄偲偡傞儌僨儖偲偐丄揤捀曽岦偺嬻偼抧暯慄嵺傛傝傕俁攞柧傞偄偲偟丄偦偺偁偄偩偼惓尫嬋慄偱寢傇儌僨儖側偳偑傛偔巊傢傟傞丅偳偪傜偺儌僨儖傕尰幚傪偐側傝扨弮壔偟偨傕偺偩丄幚梡揑偵偼巊偊傞傕偺偲傒側偝傟丄傂傠偔巊傢傟偰偄傞丅
傓偢偐偟偄偺偼丄岝尮偺柧傞偝偺愨懳揑側抣傪寛傔傞偙偲偱偁傞丅抧昞偵撏偔岝偼丄戝婥拞偺悈忲婥傗塤丄偦偺傎偐偺棻巕偺検側偳偺晄妋幚側曄摦梫場偵埶懚偟偰偍傝丄堦斒惈偺偁傞幃偱昞尰偡傞偙偲偑偱偒側偄丅掕検揑偵岝崌惉惗嶻傪尒愊傕傝偨偄応崌偵偼丄揟宆揑側惏偺擔偲偔傕傝偺擔偺柧傞偝偺帪娫曄壔傪側傫傜偐偺幃偱梌偊傞側傝丄岝僙儞僒乕偱楢懕應掕偟偨惗偺僨乕僞傪梌偊傞側偳偟偰寁嶼偡傞偙偲偑懡偄丅偩傟偱傕棙梡偱偒傞擭娫昗弨岝娐嫬僨乕僞儀乕僗偺傛偆側傕偺偑奺抧偵惍旛偝傟傟偽棙梡壙抣偼崅偄偼偢偱偁傞丅岝崌惉偺岝埶懚惈偼旕慄宍偩偐傜丄寧暿擔幩検偲偄偭偨僨乕僞偱偼晄廫暘偱丄弖娫抣偺僞僀儉僔儕乕僘偺傛偆側傕偺偑朷傑偟偄丅
偲偙傠偱丄怉暔孮棊偺壓偺傎偆偱惗妶偟偰偄傞怉暔偵偲偭偰偼丄忋憌偺怉暔偨偪偑嬻偐傜偺岝傪偝偊偓傞僼傿儖僞乕偲側傞丅怷椦偺壓偱偼傢偢偐侾乣悢僷乕僙儞僩偺岝偟偐摟夁偟偰偙側偄偙偲傕傑傟偱偼側偄丅偙偺岠壥傪昡壙偡傞偺偵偟偽偟偽巊傢傟傞偺偑丄夋妏偑180搙偺嫑娽儗儞僘傪巊偭偰嶣塭偟偨抧暯慄偐傜偆偊偺敿媴慡懱偺幨恀丄慡揤幨恀偱偁傞丅嶣塭偝傟偨慡揤偺僀儊乕僕傪揔摉側悢乮悢昐屄側偳乯偺彫嬫夋偵暘妱偟偰丄奺嬫夋偺嬻寗棪亖嬻偺尒偊傞斾棪傪撉傒庢傞丅偙傟偲偦偺曽岦偺嬻偺柧傞偝偲傪妡偗崌傢偣傞偙偲偱丄椦偺壓偵摓払偡傞岝偺曽岦暘晍傪抦傞偙偲偑偱偒傞丅
婡擻偲娭學偑偁傞峔憿側傜偽丄帺慠慖戰偺寢壥偲偟偰偺'崌棟惈'偑尒偄偩偝傟偰摉慠偱偁傞丅儅僋儘側峔憿偺側偐偵傕丄岝傪庴偗庢傞偲偄偆婡擻偵徠傜偟偰峫偊傞偲側傞傎偳偆傑偔偱偒偰偄傞偲擺摼偡傞傛偆側僷僞乕儞偑偄傠偄傠尒偮偐傞丅
偨偲偊偽丄傛偔惏傟偨壞偺擔偺恀偭拫娫丅懢梲偐傜偺捈幩岝偼偠傝偠傝偲擬偄丅慜偵弎傋偨傛偆偵丄嫮偡偓傞岝偼怉暔偵偲偭偰傕偐偊偭偰奞偵側傞偙偲偑偁傞丅梩柺偵偁偨傞岝偺嫮偝偼丄梩柺偺妏搙傪曄偊傞偙偲偱挷愡偱偒傞丅偁偊偰嫮岝傪旔偗偰偄傞偲夝庍偱偒傞傛偆側梩偺偮偗偐偨傪懡偔偺怉暔偱尒傞偙偲偑偱偒傞丅
儅僀僤儖僥儞僫儞僔儑僂傪嵽椏偵偟偨尋媶傪徯夘偟傛偆(5)丅僒僩僀儌壢偺懡擭惗憪杮偱偁傞偙偺怉暔偼丄廫悢枃偺彫梩偲傛偽傟傞晹暘偵暘偐傟偨彾忬暋梩傪偮偹偵堦枃偩偗偮偗傞乮恾係乯丅梩偺柺傪抧柺偐傜帩偪忋偘偰偄傞宻偺傛偆側晹暘偼梩暱偱偁傞丅梩傪偮偄偗偰偄傞偺偼弔愭偐傜壞偺巒傔傑偱偺悢儢寧偺傒偱丄偁偲偼搚偺拞偺僀儌偺偐偨偪偱媥柊偟偰偄傞丅
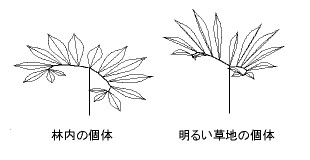
| 恾係丂 寁嶼婡偺拞偵嵞峔惉偟偨儅僀僤儖僥儞僫儞僔儑僂偺儌僨儖丏丂 暯柺揑側梩傪帩偮椦偺壓偺屄懱乮嵍乯偲丄彫梩偑婲偒忋偑傝愜傟偨偨傑偭偨柧傞偄憪抧偺屄懱乮塃乯丅梩偼侾俆枃偺彫梩傪帩偮彾忬暋梩丅 |
儅僀僤儖僥儞僫儞僔儑僂偼丄憪抧傗偁傑傝埫偔側偄椦偺壓偵惗堢偟偰偄傞丅椦撪偵惗堢偟偰偄傞屄懱偼梩慡懱偑暯柺揑偵峀偑偭偰偄傞丅偦偺柺偺岦偐偆曽岦偼偐側傜偢偟傕恀忋偱偼側偄丅 忋曽傪偍偍偆栘乆偺栁傝偐偨傗嬻寗偺暘晍偵偍偆偠偰丄傕偭偲傕擖幩岝偑戝偒偔側傞傛偆側曽岦偵岦偄偰偄傞丅岝偑晄懌偟偑偪側娐嫬偺側偐偱偡偙偟偱傕懡偔偺岝傪庴偗傜傟傞宍傪偲偭偰偄傞偺偩丅
偙傟偵懳偟偰丄椦偺奜偺柧傞偄娐嫬偵惗堢偟偰偄傞屄懱偱偼丄堦枃堦枃偺彫梩偑悈暯柺偐傜婲偒忋偑偭偰幬傔偵側偭偰偄傞偺偵壛偊丄偦傟偧傟偺彫梩偑拞墰偺柆傪拞怱偵偟偰愜傟偨偨傑偭偰偄傞乮恾係乯丅偦偺偨傔丄偍偍偔偺彫梩偺柺偼40乣50搙傕悈暯柺偐傜孹偄偰偄傞丅偙偺傛偆側宍懺偼丄擔拞偵忋曽偐傜偦偦偖懢梲偐傜偺嫮岝傪旔偗傞偺偵栶棫偭偰偄傞傛偆偵尒偊傞丅寁嶼婡偺側偐偵儅僀僤儖僥儞僫儞僔儑僂偺峔憿傪嵞峔惉偟偰寁嶼傪峴偭偨寢壥丄彫梩偺婲偒忋偑傝偲愜傟偨偨傑傝偵傛偭偰丄嫮岝慾奞傪傂偒偍偙偡傛偆側嫮偄岝偑偁偨傞帪娫偑偄偪偠傞偟偔抁偔側傞偙偲偑妋偐傔傜傟偨丅傑偨丄恓嬥傪巊偭偰梩柺傪悈暯偵屌掕偟偨偲偙傠丄惏傟偨擔偺擔拞偺岝崌惉懍搙偼帺慠偵愜傟偨偨傑偭偰偄傞屄懱偺敿暘埲壓偵傑偱掅壓偟偨丅梩偺柺傪孹偗傞峔憿偼丄偨偟偐偵嫮岝偺奞傪旔偗傞偺偵峷專偟偰偄傞丅
栰奜偺怉暔偺梩偺孹偒傪拲堄偟偰尒偰傒傞偲丄岝偑晄懌偟偦偆側偲偙傠偱偼柧傞偄曽偵岦偄偨梩丄嫮偄岝偑摉偨傝偦偆側偲偙傠偱偼悈暯柺偐傜孹偄偨梩丄偲偄偆僷僞乕儞傪偄偔傜傕尒偮偗傞偙偲偑偱偒傞丅儅僀僤儖僥儞僫儞僔儑僂偼梩傪堦枃偟偐帩偭偰偄側偄偑丄懡偔偺梩傪偮偗傞怉暔偺応崌偵偼丄嫮偄岝偵偝傜偝傟傞忋曽偺梩偑孹偄偰偄傟偽嫮岝偺奞傪夞旔偡傞偲摨帪偵傛傝壓曽偺梩偵傕岝傪暘偗傞偙偲偱慡懱偲偟偰偺岝崌惉惗嶻傪戝偒偔偡傞偙偲傕偱偒傞偼偢偱偁傞丅
偝偰丄堦杮偺宻偵暋悢偺梩偑偮偄偰偄傞偲偒丄梩偺偁偄偩偵偼擔堿偵偡傞傕偺丄偝傟傞傕偺偑弌偰偔傞丅偙偺憡屳旐堿偺偍偙傝偐偨偼丄偝傑偞傑側宍懺揑梫慺偵埶懚偡傞丅傑偢丄梩偺悢偑懡偄傎偳旐堿偑偍偙傝傗偡偄偙偲偼偡偖暘偐傞丅岝晄懌偺娐嫬偱偼梩傪偮偗偡偓偰偼偄偗側偄丅偦傟偱傕丄梩偲梩偺偁偄偩偺娫妘傪傂傠偔偁偗傟偽旐堿傪尭傜偡偙偲偑偱偒傞丅偟偐偟丄梩偺娫妘傪傂傠偘傞偨傔偵宻偵帒尮傪巊偊偽偦偺傇傫梩傪嶌傞偨傔偺帒尮偑尭偭偰偟傑偆丅椉幰偺僶儔儞僗傪掕検揑偵峫椂偡傞偙偲側偟偵丄偐傫偨傫偵懝摼傪榑偢傞偙偲偼偱偒側偄丅
梩偺宍傕娭學偡傞丅摨偠柺愊偺梩偱偁偭偰傕丄僼僉偺傛偆側娵偄梩偑宻偵枾拝偟偰偄傞応崌偲丄挿偄梩暱偑偁傞応崌傗梩偑嵶挿偄応崌偲偱偼丄憡屳旐堿偺掱搙偼戝偒偔偙偲側傞丅傛傝峀偄嬻娫傪巊偭偰梩傪揥奐偡傟偽丄偦傟偩偗梩摨巑偺旐堿偼偍偙傝偵偔偔側傞(6)丅傑偨丄抁偄宻偵懡偔偺梩偑偮偄偰偄傞応崌偵忋偺梩偲壓偺梩偺廳側傝偁偄傪旔偗傞偆偊偱偼丄梩偺偁偄偩偺梩暱偺挿偝偺堘偄傗丄悈暯柺偐傜偺孹偒偖偁偄偺堘偄丄堦枃偺梩傪弌偡曽岦偲師偵弌偡梩偺曽岦偲偺側偡妏搙側偳傕廳梫偩(7)丅
偡偙偟偱傕懡偔偺岝傪庴偗傛偆偲偄偆峔憿傗丄嫮偄岝傪旔偗偰偄傞峔憿側偳丄尒偮偗偰傒傛偆偲偄偆栚偱栰奜偺怉暔傪尒搉偡偲丄偙偺傎偐偵傕偨偔偝傫偺乽側傞傎偳乿傪敪尒偡傞偙偲偑偱偒傞(8)丅
偝偰丄偙偙傑偱偼怉暔偺惷揑側峔憿偲偦偺庴岝婡擻偲偺娭學傪峫偊偰偒偨丅偠偭偝偄偺怉暔偼丄岝崌惉嶻暔傪棙梡偟偰宻傪怢偽偟怴偨偵梩傪偮偗傞丅偁傜偨側婍姱偺敪惗偼僶僀僆儅僗偺憹壛偲偄偆堄枴偱偺惉挿偱傕偁傞偑丄摨帪偵岝妉摼峔憿偺奼戝偲尒傞偙偲偑偱偒傞丅怉暔偑惉挿偟偰偄偔偲偒丄婛懚偺峔憿偺偳偙偐傜偳傟偩偗怴偟偄宻傪怢偽偟丄偦偙偵梩傪偮偗傞偐丄偦偺僟僀僫儈僢僋側僾儘僙僗偑偳偺傛偆偵惂屼偝傟丄偦傟偑岝妉摼峔憿偺婡擻偲偳偺傛偆偵偐偐傢偭偰偄傞偺偐丅怉暔偺惉挿傪傛傝怺偔棟夝偡傞偨傔偵傕丄傑偨丄怉暔摨巑偺岝傪傔偖傞嫞憟娭學傪儊僇僯僗僥傿僢僋偵棟夝偡傞偨傔偵傕丄傂偲偮偺帪娫抐柺偩偗傪尒傞偺偱偼側偔惉挿偺夁掱慡懱傪尒傞偙偲偑昁梫偩丅
宻偑堦杮偩偗偺扨幉揑側峔憿偟偐嶌傜側偄怉暔偱偼丄宻傪怢偽偡偐怢偽偝側偄偐偖傜偄偟偐岺晇偺偟傛偆偑側偄丅偟偐偟丄暘巬峔憿傪帩偮怉暔偱偼峔憿偺壜慪惈偼偼傞偐偵崅偄丅暘巬偺僷僞乕儞偼丄怉暔偺俁師尦峔憿傪寛傔傞廳梫側梫場偱偁傞丅
宻偺巬暘偐傟偺偟偐偨偼偁傜偐偠傔僾儘僌儔儉偝傟偰偄傞晹暘傕戝偒偄偑丄懡偔偺怉暔偼丄娐嫬忦審偵墳偠偰僼儗僉僔僽儖偵宍懺傪嶌偭偰偄偔擻椡傪帩偭偰偄傞丅庽栘偱偼丄柧傞偄偲偙傠偱偼崅偄昿搙偱巬暘偐傟偟偰枾側梩孮傪嶌傞偺偵懳偟丄埫偄偲偙傠偱偼暘巬偺昿搙偑掅偔梩偺暘晍枾搙偑彫偝偔側傝偑偪偩丅岝娐嫬偵埶懚偟偨暘巬昿搙偺曄壔偼丄屄懱偺偁偄偩偩偗偱側偔丄堦屄懱偺拞偺柧傞偄晹暘偲埫偄晹暘偺偁偄偩偱傕偍偙傞丅 Koike(9)偼丄庽姤乮梩傪偮偗偨巬偑廤傑偭偨晹暘乯撪偺柧傞偄偲偙傠偺巬偼傛傝懡偔偺丄偐偮挿偄巕巬傪嶌傞偺偵懳偟丄埫偄偲偙傠偺巬偼彫悢偺抁偄巕巬傪嶌傞偙偲傪帵偟偨丅
Takenaka (10) 偼丄偦偺傛偆側巕巬偺敪惗悢偺岝娐嫬埶懚惈偲丄夁搙偵斴堿偝傟偨巬偑巰朣偡傞偙偲傪壖掕偟偰丄庽姤偺敪払僾儘僙僗偺悢抣僔儈儏儗乕僔儑儞傪偍偙側偭偨丅偦傟偧傟偺巬偺岝娐嫬偼丄屄懱偺嬻娫峔憿傪傕偲偵愭偵徯夘偟偨乽尒忋偘朄乿傪巊偭偰寁嶼偟偰偄傞丅惉挿偵偲傕側偭偰巬偑崿傒崌偭偰偔傞偲丄偟偩偄偵旐堿偝傟傞巬偑懡偔側傞丅偦偺寢壥丄巕巬偺娫堷偒傗怴偟偄巬偺敪惗偺梷惂偑偍偙偭偰巬偺夁搙偺崿傒崌偄偵僽儗乕僉偑偐偐傞丅傑偨丄壗杮傕偺栘傪側傜傋偰壖憐揑側椦傪嶌偭偰僔儈儏儗乕僔儑儞傪偍偙側偆偲丄椦偺拞墰晹偺屄懱偼擔堿偵側偭偨壓巬傪棊偲偟偰偄偒丄巬偺側偄姴偺偆偊偵庽姤偑偺偭偨峔憿偑偁傜傢傟傞丅傑偨丄椦偺傊傝偺屄懱偱偼丄柧傞偄傎偆偵岦偐偆巬偑傛偔怢挿偟暘巬偡傞寢壥丄椦奜偺柧傞偄曽岦偵敪払偟偨庽姤偑偱偒傞乮恾俆乯丅偳偪傜傕幚嵺偺椦偱尒傜傟傞庽宍偱偁傝丄傑偨丄屄懱慡懱偺岝妉摼偺揰偐傜尒偰崌棟揑側傕偺偩丅
| 恾俆丂 廃埻偺屄懱偲偺憡屳嶌梡偵傛傞儌僨儖庽栘偺庽宍偺壜慪揑側曄壔丏丂 嵍偐傜塃傊弴偵丄堦杮偩偗扨撈偱惉挿偝偣偨儌僨儖庽栘丄25杮傪傑偲傔偰堢偰偨儌僨儖庽栘偺椦丄椦偺拞墰偵埵抲偡傞屄懱丄椦偺墢偺屄懱丅椦墢偺屄懱偱偼壓偺巬偑墵惙偵怢揥偟偰偄傞塃懁偑椦偺奜偺曽岦丄壓巬偑棊偪偰偄傞嵍懁偑椦偺撪懁偺曽岦丅乮暥專(10)傪夵曄乯 |
偙偺僔儈儏儗乕僔儑儞偱偼丄栘偺抧忋晹偺儅僋儘側峔憿傪惓柺偐傜埖偄丄嬊強揑側岝娐嫬偵墳偠偨惉挿僾儘僙僗偺僐儞僩儘乕儖傪壖掕偡傞偙偲偱丄屄懱慡懱偺峔憿偺壜慪惈傗栘乆偺偁偄偩偺憡屳嶌梡傑偱昞尰偟摼偨丅偙偺偙偲偼丄慺夁掱偐傜壖憐怉暔傪慻傒忋偘偰慻傒崬傫偩慺夁掱偺堄枴傪峫偊傛偆偲偄偆傾僾儘乕僠偵偠傘偆傇傫惉嶼偑偁傞偙偲傕帵嵈偟偰偄傞偲巚偆丅
杮峞偺偼偠傔偵丄傢傟傢傟偑崱傑偱偵摼偨怉暔偑惗偒傞巇慻傒偺抦幆傪摑崌偟偰壖憐揑側儌僨儖怉暔傪嶌傞偙偲傪採埬偟偨丅偦偟偰丄偦偺晄壜寚偺峔惉梫慺偲偟偰丄怉暔偺儅僋儘側俁師尦峔憿偲奜奅偐傜岝帒尮傪庢傝崬傓婡擻偲偺娭學偵偮偄偰徯夘偟偰偒偨丅
儌僨儖偼尰幚偺怉暔偲偼堘偆丄儌僨儖傪嶌偭偨偲偙傠偱惗偒暔偑暘偐傞傢偗偱偼側偄丄偲偄偭偨斸敾傕偁傞偩傠偆丅儌僨儖偑惗偒暔偺戙懼暔偱側偄偺偼摉慠偱偁傞丅偟偐偟丄屄乆偺抦幆傪僶儔僶儔偵暲傋偰抲偔偩偗偱偼丄傕偭偲暘偐傜側偄丅屄懱傪嵞峔惉偟傛偆偵傕丄晄廫暘側抦幆偟偐側偄晹暘傕傑傑偁傞偩傠偆偑丄偦傟偼偦傟偱偟偐偨偑側偄丅傓偟傠丄儌僨儖傪嶌傞偙偲偱偳傫側抦幆偑寚偗偰偄傞偺偐傕偼偭偒傝偟偰偔傞偩傠偆丅
儌僨儖怉暔偼丄傢傟傢傟偺尰嵼偺抦幆傪摑崌偟偦偺慡懱憸傪攃埇偡傞偨傔偺摴嬶偲偟偰丄愊嬌揑偵棙梡偟偰偄偔壙抣偑偁傞偲巚偆丅屄乆偺抦幆偼丄偦偺傛偆側宍偱摑崌偝傟偨偲偒偵偙偦丄尰幚偺偝傑偞傑側栤偄偵懳偟偰掕検揑側摎偊傪梌偊傞傕偺偲偟偰惗偐偝傟偰偄偔偵堘偄側偄丅