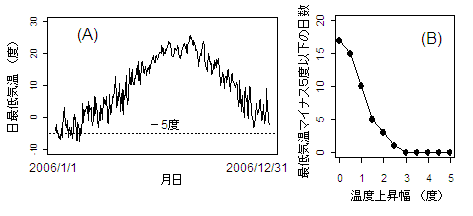
| 図1 茨城県つくば市での、毎日の最低気温(左)と、0度〜5度温暖化したと仮定して計算した、マイナス5度以下になる日数(右)。2006年のアメダスの測定データをもとに計算した。 |
この文章は, 日立環境財団発行の 「季刊・環境研究」に掲載する記事の原稿として執筆したものです. 最初の入稿段階のもので,印刷されるものと同一とは限りません.
※原稿依頼時にいただいたお題は「地球温暖化と生物多様性」でしたが, 生物多様性という語はもともと科学的に定義された概念というよりは 保全のためのキャッチコピー的な言葉として作られたと理解しています. 本稿ではあえてこの語を使わずに,生態学的な観点から地球温暖化を含む気候変動の影響を論じてみました.
(2008-01-26)
この原稿を書いている今は冬の最中である。北関東のつくばの冬は寒い。早朝の庭は数センチの霜柱が土を持ち上げている。暖かい春が待ち遠しい。庭の植物もじっと春を待っている。南の植物であるハイビスカスはこの冬が越せない。そんな寒さの中でもスイセンは花を咲かせている。種類によってずいぶん違うものである。
世の中では温暖化が進んでいる。人間は、石炭や石油を燃やして多量の二酸化炭素を大気中に送り込んだ。それが原因で地球は温室のように冷えにくくなり、地球全体の平均気温が少しずつ上がっている。気候変動に関する政府間パネルの2007年の報告書によれば、温暖化が進んでいることはもはや疑いがなく、その原因が人間活動である可能性はかなり高く、今後も温暖化傾向が続くことはほぼ確実であるという(1)。最近100年のあいだに地球の平均気温は0.7度あまり上昇した。温暖化の傾向はとくに最近30年間で顕著である。
地球の歴史のなかで気候の変動は繰り返されてきた。そのなかには、数十年のあいだに数度も温度が上下するという劇的な変化もあったと考えられている。それらの比べ、いま進行中の地球温暖化が特異なのは、たったひとつの種である人間の活動が急激な変化をもたらしていることである。前例がない大事件であり、我々はみずからの振る舞いが引き起こしたことに責任を感じる必要がある。
温室効果気体の濃度をかりに現在の状態に保つことができたとしても(それはほぼ無理なのだが)、今後20年間のあいだにさらに0.2度程度の温度上昇が予想されている。実際には、温室効果気体の濃度はさらに上昇を続け、温度の上昇もさらに大きいものになると予想されている。現在の気候条件を前提に発展してきた人間の社会にとって、温暖化の進行は、暖かくなってうれしいと喜んでいられるようなものではない。さまざまな不都合が生じるだろう。
生き物の自然はどうだろうか。北海道と沖縄、あるいは北アルプスの山頂近くと麓とを比べてみれば、それぞれに特徴のある自然が見られる。涼しいところは涼しいなりの、暖かいところには暖かいなりの自然がある。生態系全体をながめても、生態系のメンバーである個々の生き物の種類に目を向けても、気候に応じた違いがある。涼しいところが急に暖かくなってしまうといろいろ不都合がありそうだ。今までつくばの庭に植えられなかった庭木が育つと喜んではいられない。
本稿では、陸上の生物を中心に、温暖化のもとでの生き物の分布域がどうなるのかを考える。さらに、われわれは何を心配しなくてはならないのか、そしてどういう対策が可能なのかにも触れたい。
日本の四季の自然のうつろいは、温度の変化に対応したものである。冬には寒さに耐える体制をとり、春になって暖かくなれば眠っていた生き物が動きだす。夏の盛りの暑さがつらい生き物は日差しを避ける。秋になれば来るべき冬への備えを始める。生物の生活は温度に強く影響される。温暖化とともに生き物の振る舞いも変化するのも当然である。
ところで、多くの生物は限られた気候条件の範囲内に分布している。亜熱帯性の気候でのみ見られる生物もいれば、永久凍土地帯や高山にだけ分布する生物もいる。乾燥したところに特有の生物もあれば湿地に特有の生き物もいる。
とくに、高緯度側(北半球なら北)や、山のどの高さまで分布するかといった、低温側の分布の限界について考えてみる。一年の平均的な気温が限界を決めているとは考えにくい。冬の最低気温の下で生き延びられるかどうか、あるいは夏の日中の温度が生育・繁殖に十分かといったように、ある時期の温度が重要な限定要因となっていることが多いだろう。庭に本来は越冬できない種類の植物を植えておくと、晩秋や冬の早朝の強い冷え込みで、急に萎れてしまうのを見ることがある。早朝の冷え込みでどこまで下がるかが、その植物が露地で越冬できるかどうかを決めている。このように生物には、これ以上寒いと(あるいは暑いと)死んでしまうという温度限界がある。そうした厳しい日があるかどうかが、空間的な分布の限界を決めていると考えられる。
このことを念頭において、一年間気温のパターンを見てみよう。図1は茨城県つくば市での、2006年一年間の毎日の最低気温のデータを示したものである。この年、最低気温がマイナス5度以下となったのは17日であった。もし気温が全体に1度だけ上がったとすると、この日数は10日となり、2度上がったら3日、3度以上あがれば1日もなくなる。マイナス5度が生きるか死ぬかの瀬戸際の生き物から見れば、この違いはとても大きい。温暖化によって温度が数度上がることがそんなに大きな影響を持つのか疑問にも感じられるかもしれないが、生物に強く影響するのは特に暑いとき、あるいは特に寒いときの気温だとすると、平均気温の数度の上昇が生物にとっては大きな変化となる。
|
| 図1 茨城県つくば市での、毎日の最低気温(左)と、0度〜5度温暖化したと仮定して計算した、マイナス5度以下になる日数(右)。2006年のアメダスの測定データをもとに計算した。 |
自然の中の生物はその生存限界ぎりぎりにまで分布しているのだろうか。植物園に植わっている植物を見ると、本来ならそこには分布していない種類を見ることがある。つくば市の平地にある植物園にはブナが植えられており、そこの環境はブナが生育可能なものであることが分かる。しかし、実際にはもっと標高の高いところに登らないと自然のブナは見られない。なぜだろうか。
重要な要因として考えられているのは、他の生き物との相互作用である。生存をおびやかすような競争相手や捕食者、寄生者、病原生物などの存在や、逆に生存に必要な生物の不在が、生物の分布範囲を制限する。潜在的な分布範囲の境界近くでは生きているだけで苦しいことが想像される。そのような状態では、他の種との相互作用の効果は大きく、それだけ競争に負けたり病気になったりしやすいだろう。
人が植えて育てれば育つ範囲と、自然のなかで他の生物に囲まれた状況で育つ範囲とが一致しないことは、生態学者が古くから認識していた現象である。自然の環境が多様な中で、ある生物が生息できる場所をあるいはニッチと呼ぶ。物理環境に関する限りは生育可能な環境は生理的ニッチ、他の生物との相互作用もあるなかで生息可能な環境は生態学的ニッチと呼ばれる。
現状の理解や将来の予測にあたって、過去に学ぶことは多い。気候変動の研究では過去のパターンの解析は欠かせない。気候変動が生物に与える影響についてもそれは同じだろう(2)。
今から約5億年前、陸上で生物が暮らすようになってから、デボン紀、ペルム紀、白亜紀の温暖な気候とそのあいだの寒冷な気候という大きな変動があり、それは生物の進化の歴史にも大きな影響を与えた。より最近の変化に注目してみると、過去数百万年は氷河期(あるいは氷河時代)であり、そのあいだはとくに寒冷な氷期と温暖な間氷期が4〜10万年の周期でおとずれている。その温度差は、5〜7度、海水面の変化は100メートルほどと推定されている。現在は間氷期であり、最後の氷期(最終氷期)が終わったのは約1万年前のことである。その時と比べ、生物の分布可能域はより高緯度に、より高標高に拡大している。なお、最終氷期といっても、もう氷河時代は終わって氷期は来ないということではない。過去の氷期のなかで最後のものという意味である。地球の気候システムが予想もつかないカタストロフィックな変化を起こさない限り、おそくとも数万年のうちにはふたたび氷期となると考えられている。
最終氷期が終わってからの1万年という年月は、生物の分布範囲の回復に十分なものなのだろうか。これは生き物の種類によりさまざまであろう。地上を主体的に動き回れる生物ならば、数千キロを移動するのにも十分過ぎる長さだ。一方、新しく定着した個体が種子をつけるまでには何十年とかかる樹木であれば、一万年でも十分ではないかもしれない。とくに、種子の散布距離が短い種類では、いまだ移動の途中であるとも言われている。
最終氷期後の生物の分布範囲の移動速度は、とくに北米とヨーロッパで詳しく研究されている。その速度は、たとえば樹木では年あたり200メートルから2キロほどと見積もられている。芽生えた個体が成熟して種子を作るまでにかかる時間を考えると、これはあまりに早いように思われる。
この速度を説明する仮説のひとつは、ごくまれに起こるきわめて長距離の種子散布が重要な役割を果たしているというものである。たまたま渡り鳥の体に付着した種子が数十キロも100キロも運ばれるといった機会が、分布の拡大に貢献しているのかもしれない。もうひとつの仮説は小さなレフュージアの効果である。寒冷化したときの避難場所はレフュージア(refugia)と呼ばれる。本来ならレフュージアとはならないはずの地域に、南向きの谷など、ごくごく局所的に寒さが緩和されたところが小さなレフュージアとなり、ここでかろうじて生き延びた小さな個体群が氷期が終わったあとに大きくなったのだが、小さなリフュージアの存在が現在では認識できないため、あたかも極めて大きな速度で北進したように見えているのだという仮説である。現在の生物の遺伝子を調べ、その空間パターンから過去の分布範囲の変遷のパターンとプロセスを推測しようとする研究が進められている。
生物の分布範囲の移動は、どのようなプロセスを経て起こるのだろうか。今の居場所は暑すぎる、あっちのほうが涼しそうだとみんなで揃って歩いていくといった単純なものではないだろう。また、生物の種類によっても異なるはずだ。長距離を自由に移動できる動物もいれば、土に根をおろしたらそれっきり移動できない植物もいる。固着性の生物でも、植物なら種子や胞子、固着性のサンゴやフジツボなら卵や幼生のときには新天地への散布が可能となる。
自分自身が動き回るにしろ、卵や種子を周囲に散布するにしろ、分布の限界近くでは、ときおり限界を越えたところへ行き着くものもいるだろう。もしそこで定着し、さらに子供を残すことに成功すれば、すなわち分布範囲を広げたということになる。逆に、行ってはみたもののたちまち死んでしまう、あるいはかろうじて生きてはいるものの子供は残せないのであれば、分布は拡大しない。
温暖化によって、たとえば北半球の生物の分布の北限がより北に移動しているということは、皆が引っ越しをしたということではなく、これまでの限界を越えたところに出かけていった個体や散布された子供が、寒さがやわらいだために定着・繁殖に成功し、さらにその北へ移動する・散布される個体が供給されるということである。
ところで、生物は一面に連続的に分布しているのではなく、あちらにひとかたまり、こちらにひとかたまりと不連続なパッチワーク状に分布していることが少なくない。個々のかたまりのレベルでみると、これまで存在していたかたまりが消失したり(局所的な絶滅)、新しいかたまりが出現したりといったことが同時に起こっている。こうした分布パターンを示す生き物の分布をマクロに見たとき、南限近くのかたまりでは局所的な絶滅が多い一方で、北限近くでは新規に発生するかたまりが多く、その一部はこれまでの分布限界を越えたところまで及ぶなら、分布は北へむけて移動していることになる。その様子を人間が捉えるには、北限で新しい定着地を見つけるだけでは不十分である。絶滅と定着のバランスを、北限近くと南限近くで比較することが必要である。
気候の変動には年ごとのランダムに見える変動もある。100年のスケールでは徐々に温暖化が進行しているとしても、暖冬もあれば冷え込みが厳しい冬もあるし、猛暑の夏もあれば冷夏もある。全国的な冷夏でコメが凶作となった1993年にも、地球レベルでは温暖化は進行していたはずである。
生き物の分布の限界も、年ごとの環境の変動によって進んだり戻ったりしていると考えられる。温暖化にともなってより高緯度へ、あるいはより高い標高へという傾向があったとしても、それにこうした一進一退の変動が重なる。数年程度の観測では長期的な傾向を確かめることはできない。分布の移動を確認するには、十分に長い期間にわたって調査すること、また、特定の地点だけで見るのではなく広い範囲での個体群の消長を調査することが必要である。
図2は、簡単なシミュレーション計算の結果である。緯度に比例して最低気温が低下する(高緯度ほど最低気温が低くなる)と仮定する。さらに、年とともに一定の速さで温暖化し、最低気温が上昇するとする。つまり、年と緯度によってその年の最低気温が決まる。ただし、実際の最低気温はランダムな変動も含むものとする。生物は、ある年の最低気温が限界温度以下になるとただちに死亡すると仮定する。寒い年には、高緯度に分布していた個体は死んでしまい、そのあと年々一定の速度で分布は回復していくとする。そのあいだに寒い年があれば、ふたたび分布は低緯度へと押し戻される。
図2(A)は、温暖化の速度がゼロ、すなわちまったく温暖化しない場合、(B)は年々温暖化する場合である。どちらの場合も、年ごとに生物の分布可能な境界(細い破線)はランダムに変動しているが、温暖化が進んでいる場合には、全体として徐々に高緯度へと移動している。一方、生物が分布している場所の限界(太線)は、特に寒い年に低位緯度へと押し戻されたあとただちに回復することはできずに徐々に高緯度へと分布を広げるから、分布限界は鋸状に変動する。それでも、温暖化が進んでいる場合には徐々に高緯度へと分布を拡大していく。 (A)図と(B)図を、100年全体で見比べればそのちがいは明らかだが、数年分だけを取り出してみれば、温暖化が進んでいない場合でも、寒い年に押し戻された分布域が徐々に回復して分布が高緯度方向へと拡大している部分があちこちにある。このような短期の調査では温暖化のために分布が拡大しているのかどうか、なんとも判断ができないことになる。十分に長期のデータを見ることの重要性が、このモデルからもよく分かる。
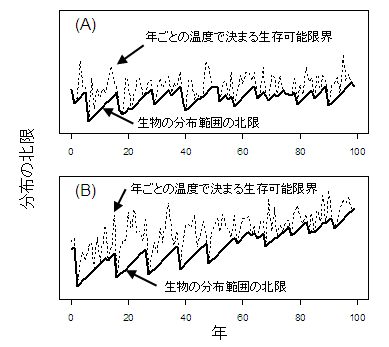
| 図2 年ごとのランダムな寒暖によって、ある種の分布限界緯度が変動する(細い破線)。生物の分布は、これを越えることはできず、寒い年にはその年の限界を越えたところに分布していた個体はすべて消滅する。そのあと、分布域の回復は瞬時には起こらず、ある一定の速度でしか高緯度へ進めないとすると、分布の境界線(太い実践)は図のように鋸状になる。これは、温暖化が進んでいなくても(A)、温暖化が一定のペースで進んでいても(B)同様である。ただし、温暖化が進む場合には、長期のトレンドを見ると徐々に高緯度へと分布が拡大している。 |
Parmesanと Yohe (2003)は、多数の生物の分布域の調査報告の中から最低でも20年以上にわたるものを選び出してその傾向を分析した(3)。これだけ長期の調査なら、短期的な変動の背景にある長期傾向が見えるはずである。分析の結果、279種のうち177種で方向性のある変化が認められ、その8割は高緯度方向への移動であった。また、ある地点での個体群の継続観察の結果では、寒冷地を本拠とするものの減少や、温暖地を本拠とするものの増加が調査対象の641種のうち36%で認められ、その反対の傾向は8%のみの種で認められた。たしかに生物の分布は高緯度へ向かって移動中である。
個々の生物の分布範囲が変化すれば、ある地点の生態系の構成メンバーも変化する。すでに簡単に触れたように、すべての生き物が歩調を揃えていっしょに動くはずはない。自由に飛び回れる鳥と、種子が芽生えて成長して親木になり、その種子がまたどこかに運ばれるまで何十年もかかる樹木とでは、移動のスピードが違って当然である。したがって、生き物たちが気候の変動に対応する過程では、現在とは異なる顔合わせと相互作用が生じるはずである(図3)。
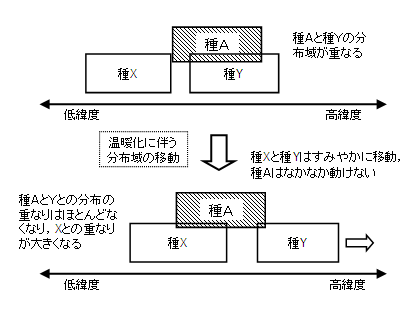
| 図3 種によって移動速度が異なることで、種間の相互作用が新たに生じたり、失われたりする。上の図では種Aは種Yと分布域が重なって相互作用が生じているが、温暖化に対応して種Yはすみやかに高緯度へと移動し、一方種Aの移動速度がきわめて遅いと、これら二種のあいだの相互作用が失われていく。もともと種Aと分布が重なっていなかった種Xが種Yと同様に高緯度へと分布を移動させると、あらたに種Aと種Xの相互作用が生じる。 |
生物のあいだの相互作用にはさまざまなものがある。同じ資源をめぐっての競争者の関係のほか、食う・食われるの関係、宿主と寄生者の関係、一方を他方が生活場所として利用する関係、植物と花粉や種子の散布者の関係などさまざまである。すべての生物は、他の生物との相互作用のなかで日々暮らしている。生態系を構成する生物が多様になれば、相互作用もまた多様となる。100種の生物がいれば約5、000通りの種間相互作用、1、000種なら500、000通りの相互作用があり得る。
生物の分布範囲を決める要因として、他の生き物との相互作用の重要性についてはすでに述べた。餌になる生物や花粉を運んでくれる生物がまわりからいなくなれば困る。競争者、捕食者や病原性生物があらたにやってくるのも迷惑である。こうした効果で、生物によっては生きつづけることが難しくなるかもしれない。あるいは、これらの逆の現象で、あらたな出会いと別れによって個体数を増やす生物もいるかもしれない。
それまで相互作用がなかった生物の初顔合わせが時として極めて深刻な影響をもたらす例は、いくらも見つけることができる。たとえば、肉食の哺乳動物がいない島に住む鳥が、捕食者から逃げる必要がないために進化の過程で飛ぶ能力を失った例がいくつも知られている。そこに人間が犬や猫を持ち込むと、飛べない鳥は逃げるすべがない。ふたたび空を飛べるように適応進化をとげるよりずっとまえに、鳥は犬や猫に食べ尽くされれてしまう。
病原生物と寄主との関係もまた同様である。かつて南アメリカのアンデス地域の人々は、ヨーロッパから持ち込まれた天然痘・はしか・腸チフスなどの病原菌により病に倒れ、人口が減少した。それまで接触したことのながった微生物に対してまったく抵抗力を持っていなかったからである。
人間の活動が人為的な国境を越えるだけでなく、海や山脈という地理的な境界をも越える。それに伴って意図的・非意図的に運びこまれた外来生物が、地域ごとの自然の構成メンバーを脅かすことはすでに繰り返し指摘されている。温暖化は、そうした脅威をよりいっそう拡大する可能性がある。
地球温暖化が深刻な問題として論じられている今現在も、大気中の二酸化炭素濃度は増え続けている。この増加を止める努力が国際的に始まっているが、スムーズには進まない。また、その努力が実を結ぶとしても、まだこのあと数十年は二酸化炭素濃度の上昇は続くだろう。もし温暖化が生き物に影響することは不可避であるならば、その影響を不可逆的なものとしない努力が、われわれができることである。そして、不可逆な変化の最たるものは種の絶滅である。
これまで、人間が生き物を絶滅においやる大きな要因には、土地の開発による生息地の破壊、捕獲・採取、生息環境の汚染、人為的な外来生物の持ち込みなどがあった。保護区の設定はこれらへのの対策として有効なものであった。保護区内の土地は開発しない、保護区内では生物を捕らない、保護区内に外来の動物を持ち込まない、保護区内を汚さないといったルールに従えば生物を守ることができる。
今、地球温暖化が生き物へのあらたな脅威となっている。これに対して保護区はどこまで有効だろうか。まず考えなくてはならないのは、ある生物を保全しようと意図して設定した保護区が、温暖化によって生育適地でなくなってしまったら、もはやその生物の保護区としては機能しないことである。その際、気候の変化そのものが生存に不適なものであるかもしれないし、気候の変化にともなってあらたに侵入してきた生物が負のインパクトを与えたり、逆に今まで餌や送粉者として利用していた生物がいなくなってしまって生存の基盤が危うくなる可能性もある。
過去の気候の変動をさぐる研究者の努力により、ダイナミックな気候の変動の歴史が見えてきた。氷期・間氷期の変動のほか、氷期のあいだに突然数十年だけ5度以上も温暖化するというダンスガード・オシュガー現象や、最終氷期が終わって現在の間氷期がはじまったあと、急激に寒さが戻った期間が1300年ほど続いたヤンガードリアス期とよばれる時があったこと、そのあと再び暖かくなる際の変化も急激で、10年ほどのあいだに何度も上昇するようなものであったことなどがわかってきた。現在、我々のまわりの自然は、こうした変化を生き延びてきたものである。しかし、だからといって現在進行中の温暖化をも生き延びられると考えるのは楽観的にすぎる。過去の気候変動でも生物の絶滅は起こった。さらに、過去と現在の大きな違いは、生物の生息地が人間によって細切れにされていることである。気候の変化に応じて分布域を移動しようにも、その前に市街地や農地が横たわっていれば、これを越えて進める生物は限られる。
従来から、細切れの保護区がそれぞれ孤立して散在しているよりも、それらのあいだを生物の行き来が可能な回廊でつなげば、地域全体での保全はより確かなものになると考えられてきた。局所的な絶滅が起こっても他の生息地からの移住で個体群が再生でくるからである。そのような保護区のネットワークは、温暖化が進む中で保全の効果をあげるうえでも有効なものであろう。現在の生息地が好ましいものでなくなっても、ネットワークをたどって新たな生息地へと移っていくことができるからである。また、ネットワークでつながれた個々の保護区も、なるべく大きな緯度方向の広がりや標高差を持つことが望ましいだろう。これからの保護区の設定と管理では、100年、200年先までの気候の変動を念頭においた、長期的な視点が欠かせない。
本稿は、地球温暖化が生物に与える影響のすべてを解説したものではない。たとえば、本稿では触れていないが、たとえば開花期の変化のように、生物が移動せずに現在の生息場所でどのような影響を受けるかというのも重要な視点である。種類によって温暖化の影響の受け方に違いがあれば、たとえば花は咲いたがそこに訪れるはずの昆虫はまだいないといったように、時間的な新たな出会いと別れを生じるかもしれない。変化した環境のもとでどれだけ遺伝的な性質が変化するのか、その可塑性は種によってどう違うのかといった問題もある。
想定される地球温暖化の影響は生態系の数、生物の種の数、種間の相互作用の数ほどある。そのすべてについて確実な予測をすることはとうていできない。自然界で何が起こっているのか、注意深く見守りながら柔軟で余裕のある保全策を講じていくことが必要だろう。