
| 図1 イチョウの短枝.3年で1センチ足らずしか伸びていない. |
第49回日本生態学会大会(2002)で,「 資源獲得戦略としての樹木の形作り − 「枝葉末節」から本質へ 」と題したシンポジウムを行いました.その内容をもとに,雑誌『生物科学』で特集 「 樹木の形作りと生き方− 「枝葉末節」から本質へ 」が組まれました. このページでは,同誌にに掲載された私の文章を同誌の了解を得て公開します. 本文中で「本誌の記事」とあるのは,この特集の他の記事のことです.
竹中明夫 (国立環境研究所)
植物は空間中に葉や根を配置して資源を獲得する.植物の形作りは動物のエサ採り行動になぞらえることができる.樹木の地上部の構造と光獲得機能についての研究は多い。しかし、それらの成果を樹木の生き方の理解へとつなげていくには、さらに乗り越えるべきハードルがいくつかある。それは、定性的議論から定量的議論へ、部分から全体へ、時間断面から木の生涯へ、そして周囲の個体との相互作用へと視点をひろげていくことである。そのためには、コンピュータを使ったアプローチも欠かすことができない.また,枝や幹の太さがどのように決まっているのかを理解することもさまざまな場面で重要となる.
動物がエサを探して行動するにはエネルギーが必要だが,エサのないところにじっとしていては飢えるばかりである.動物の採餌行動は,かけるコストと得られるエサとの兼ね合いのなかで進化してきた.いっぽう,動物のように動き回ることができない植物は、空間中に葉や根を配置して光や水を獲得する.広く枝と葉をひろげれば光を受ける面積は大きくなる.周囲の植物よりも高く伸びれば,日陰にされることなく光を受けることができる.しかし,広がるにしろ高く伸びるにしろ,体を作るための材料とエネルギーを必要とする.いわば成長のコストである.このコストは枝を細くすれば減らすことができる.けれども,自分自身を支えることができなかったり,ちょっとした力で折れてしまったりするほど細くては困る.また,葉からの蒸散をまかなう水の通り道の確保も必要だが,細すぎる枝では水の供給が追いつかないかもしれない.となると,いくらでも細くしてよいというわけには行かない.資源獲得のために広い空間に根や葉を置きたい,でもそのためにはコストがかかるというジレンマのなかで樹木は進化してきた.
これまで,樹木のマクロな形の機能的な側面の研究は,個体の高さ,樹冠(葉や枝がしげっている部分)の投影面積,葉面積の高さ分布などに注目して行われてきた.しかし近年,地上部の形のパターンとそれが作られるプロセスにより深く踏み込んだ研究が盛んになっている.個々の枝での茎と葉のバランス,分枝パターン,枝の伸びる方向などを調べ,その資源獲得上の意義を考える研究である.
そうした研究の成果を,生態学的な文脈のなかでの樹木の生き方の理解へとつなげるにはどうしたらよいのか.最近の研究の蓄積は,はたして本質の理解へとつながるものなのか.本稿では,木の形に見られるパターンとその機能的な意味について,これまでの研究を簡単に紹介しながら今後の研究の方向を考えてみる.
一本の茎とそこについている葉をひとまとめにしてシュートと呼ぶ.どこからどこまでを一本の茎と呼ぶのかはっきりした定義はない.以下では一年の生育期間中に伸長した部分をシュートの単位として考えることにする.どんな巨木も,年々伸びるシュートが集まってできあがる.木の形作りのプロセスは,シュートの生成,伸長,肥大,そして死亡のプロセスに還元して考えることができる.
おなじ種類の木が長枝と短枝というふたとおりのシュートをつくることがよくある.長枝は,われわれがふつうに枝だと認識するような,長さが数センチ〜数十センチと細長く伸びた茎に葉がついたものである.いっぽう短枝は,枝が数ミリ〜1センチ程度とほんのわずかしか伸びない,ようするに寸詰まりの枝である.その短い茎に何枚も葉をつけているものも多い.イチョウ(図1)やカラマツなどは典型的な例だが,そのほかにも多くの種類が長枝と短枝を作ることが知られている.
|
| 図1 イチョウの短枝.3年で1センチ足らずしか伸びていない. |
機能面から見ると,長枝はあらたな空間へと伸びていく伸長指向のシュート,短枝はその場で光を受ける葉の足場となる葉面積展開指向のシュートと見ることができそうである.はっきりした短枝を作る樹種にかぎらず,伸長指向のシュートと葉面積展開指向のシュートを作りわけている樹種は多い.たとえばTAKENAKA (1997)は,短枝の分化がほとんど見られない常緑広葉樹8種を調べた結果,長いシュートは長さのわりに葉面積が大きくない伸長指向シュート,短いシュートは短いながらも何枚も葉をつけている葉面積展開指向シュート,という傾向を見いだした.また,YAGI & KIKUZAWA (1999)は短枝の分化がはっきりした種,そうでない種あわせて8種類を調べ、そのうち7種類で同様の傾向を見いだした.形態的にはっきりと短枝が分化した種に限らずに、伸長指向のシュートと葉面積展開指向のシュートをあわせ持つのはかなり普遍的なようだ.
この現象にはどんな意味があるのだろうか.ひとつの仮説として、枝を伸ばしてあらたな空間に進出する役割とその場で葉を広げて光を受ける役割をシュート間で分担すると,支持器官にかけるコストが少なくてするのではないかと考えられている。この仮説を検証するには,分業をする場合としない場合それぞれについて、構造作りのコストと受けられる光の量を調べればよいはずである.一本のシュートだけで考えれば短枝のほうが効率がよいに決まっているが、短枝ばかり作っていては,狭い空間に葉が密集してしまって具合が悪い。個体全体として考えないといけない. シュートから個体へ、すなわち部分から全体へと研究を進める必要がある.
しかし、これは口で言うほど簡単ではない.ある分枝構造をもつ木の各部はどれだけの強度を持たなければならず、そのためにはどれだけ太くする必要があるかを定量的に評価しないと、コストを決められないからだ.じつのところ,枝や幹の肥大成長についは、記載的な研究は多いものの,力学的に考えてどれけの太さが必要かを押さえた研究はごく少ない(FORD et al. 1990など).とはいえこれを克服しないと,ある構造を持つことが個体全体でどれだけのメリットがあるのかを定量的に示すことはできない.今後の課題である.
なお,本特集中の鈴木氏の記事では,シュートの構造と機能を詳しく解析した例が紹介されている.また,シュート作りと肥大成長の関係や,繁殖への投資との兼ね合いについても触れられている.
図2はモモタマナ(Terminalia catappa)という熱帯を中心に分布している木である。この木は、側枝が横に広がってできる円盤が幹を軸として重なった構造を作る。ちょうど五重塔のような印象の構造だ。こうした樹形はモモタマナにかぎらず,ミズキ属の樹木など,少なくない樹種で見られる.

| 図2 モモタマナの樹形.数本の側枝が輪生状について円盤を作っている. |
平面状にひろがる側枝をよく見ると,規則的な分枝パターンに気付く.アメリカの研究者J. FISHERは、一定の分枝パターンの繰り返しでモモタマナの樹形ができあがることを目に見える形で示したいと考えた。そのための道具として,本多久夫のモデル(HONDA 1971)に注目した.このモデルでは,木を線分の組み合わで表現する.一本一本の線分が枝に相当する.親枝の先から2本の新しい子枝を伸ばすプロセスを繰り返して樹木状の分枝構造が作られる.子枝と親枝のなす角度はパラメータとして与える.また,それぞれの子枝の長さは親枝の長さに比例定数をかけて決める.分枝角度や親枝と子枝の長さの関係を変えると,さまざまな3次元構造ができてくる。モモタマナにあわせて,中心で直立する幹とそこから分枝する側枝とを区別するなど多少の拡張をしたうえでてパラメータの値を調節しシミュレーションしてみると、とてもそれらしい樹形が再現できた(FISHER & HONDA 1977).
本多氏のモデルが発表された30年前とくらべれば,コンピュータの計算速度は文字どおりケタ違いに速くなったし,個人の机のうえに自由に使えるコンピュータが乗っている.その気になればだれでも手軽にシミュレーション計算を試みることができる。図3に示したのは,私の自作プログラムで描いたさまざまな樹状の分枝構造である.
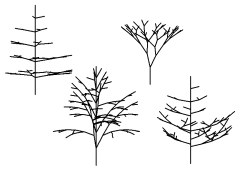
| 図3 コンピュータのなかで分枝プロセスを再現して描いた樹状の分枝構造.分枝の角度や親枝と子枝の長さの比率などを変えると,全体の形もさまざまに姿をかえる. |
コンピュータで樹形を再現することは楽しいが,研究のうえでどのような意味があるのだろうか.利点のひとつは、3次元の分枝構造というややこしいものが、ごく少数のパラメータで記述できることである.構造自体を記述するかわりに,少数のパラメータを含む生成ルールで記述するのである. そのルールからできあがる形はいつでもコンピュータが描いてくれる.
もうひとつの利点は,分枝ルールのパラメータの値を変えていろいろな仮想実験ができることである.木の実物の形態を自由に変えることはできない.むりにやろうとすれば非常にたいへんであろうことは,盆栽作りの手間を見ても分かる.HONDA & FISHER (1978) は,コンピュータ中の仮想樹木の利点を生かして,分枝パターンと受光効率の関係を調べた。Terminalia の葉は、枝の先や、分岐点に残っている短枝の先端にまとまって輪生状につく。だから、側枝全体を上から見ると,緑の小さい円盤がたくさん集まったような構造になる。この小円盤はなるべくお互いに重なり合わないほうが、側枝全体が受ける光の量は多くなってつごうがいいだろう.枝の長さをのばせば重なりを避けられるが,それには枝作りのコストがかかる。一方,分枝の角度を変えるぐらいならたいしてコストは変わらないだろう.分枝角度をいろいろ変えてみて側枝全体の投影面積を調べたところ,子枝の分枝角度の組み合わせによって、側枝の投影面積(すわわち受光面積)は変化した.そしてモモタマナの枝の分枝角度は受光面積が極大となる値にごく近いことものだった.この研究は,モモタマナの分枝パターンの合理性を示しただけではなく,コンピュータの中に樹形を再現することが構造と機能を研究するうえで強力な道具になりそうだということも示している.
ところで,HONDA & FISHERはは側枝のなかでの葉群の相互被陰を評価しているが,側枝群がつくる円盤同士の日陰にする,されるの関係は考えておらず,側枝から個体全体へとスケールアップすることは今後の課題として残されている.個体レベルでの受光効率は,円盤の間の距離に依存しそうである.円盤の間の幹の長さが短ければ,下の円盤にとって上の円盤はとてもじゃまである.一方,幹を伸ばして円盤の間隔を長くすれば相互被陰はすくなるなるが,幹を伸ばすにはコストがかかる.さらに,側枝の上にどれだけ密に葉を展開するかも個体の受光量に大きく影響しそうだ.
HONDA & FISHER (1978) らの研究から4半世紀ほどたったいま,コンピュータのなかに仮想植物を作り,その構造を操作しながら受光機能を評価することははるかに容易になった(たとえばMURAOKA et al. 2000, TAKENAKA et al. 2001).しかし,構造をつくるためのコストをあわせて考えないと,個体全体の効率を評価することはできない.前節の場合と同様の課題である.
離れたところからは緑のかたまりに見える木でも,その真下に立って見上げてみるとそこにはすかすかの暗い空間があり,葉はほとんどない。また、林のなかの木は幹の上方に葉が集中していて,下のほうには枝がも葉もあまり見られない.林の縁の木では、林の外側にむいた明るい側だけ地面近くまで葉をついてる。このように光が豊富なところに葉が集まっている構造は,光を獲得する機能の点からはつごうがよさそうである.
ところで,木のあちこちの枝を見比べると,明るいところの枝からはたくさん子枝が分枝しているが,暗いところの枝ではほとんど分枝していない.もっと暗いところには枯れた枝もたくさん見つかる. TAKENAKA (1994)は,このような光環境に依存した枝の生き死にや枝分かれの多い少ないによって明るいところに葉が集中した構造ができるのだろうと考えた.このメカニズムでうまく樹冠の形成プロセスを再現できるか、見やすい形で確かめるために,HONDA(1971)の3次元樹形モデルを拡張して,末端の枝には葉のかたまりを持たせ,それらの相互の日陰にする,されるの関係を計算した.そして,枝ごとの光環境におうじて生死や小枝の数を変えてみた.すると, 明るいところの枝はよく分枝するいっぽうで暗いとこの枝は枯れ落ちた結果,樹冠全体としては明るいところに葉を集中させた構造ができあがった.さらに,複数の個体を並べて育てると,お互いに日陰にしあう部分の枝が枯れ落ちることで,林のなかの木の形まで再現できた(図4).

| 図4 子枝の数や枝の生き死にが,枝の置かれた光微環境に依存すると仮定して,コンピュータのなかで育てた仮想森林.森林の中央部の木では下枝が光不足のために枯れ落ち(b),林縁に位置する個体では林外の明るい側に面した側だけ下枝が発達する. (Journal of Plant Research 107:321-330. より引用) |
このモデルでは,ある時点での光の空間分布が次の時点での木の形作りに影響し,作られた木の形がさらに光の分布に影響しというように,木の資源獲得構造と,光という資源の空間分布とのダイナミックな相互作用を再現している.となりあう木々は,光の空間分布を介して相互作用し,お互いの形作りに影響しあう.個体間の相互作用と木の形作りの関係については、本特集の梅木氏,隅田氏の記事でも触れられている.
このように,枝の成長や生き死にと光資源の獲得機能との関係を表現できるならば, 樹種による成長パターンの違いを,それぞれの生き方と結びつけていくことができそうである.しかし,なるべく高く,広く伸びたほうが光を受けるには有利なのは自明である.無制限な拡大を制限しているのは,高くなるにも広がるにも必要な成長のコスト,すなわち枝を伸ばしたり太らせたりするのに必要な材料やエネルギーである.そのコストを考えることなしに,どれが有利だとか不利だとか議論することはできない. 形作りのコストと利得の評価にまで踏み込んでいくことが今後の重要な課題だが,ここでもふたたび肥大成長の必要量を見積もることが必要となる.
前節で紹介したモデルでは、枝の振る舞いはそれぞれがどれだけ光を受けているかで決まり,個体内のほかの枝は無関係だと仮定している. ところで,植物体を構成するユニットは,それぞれがかなり自律的に生きているという見方がある(SPRUGEL 1991)が,もちろん完全に独立なわけではない.個体内でいろいろな相互作用があるし,ものの流れもある.本特集の長谷川氏の記事では,樹木の体内での光合成産物の流れとその研究方法について解説されている.
シュート間の相互作用の例をひとつ簡単に紹介する。たいていの場合,一本の木の中でも日当たりがよい枝と日陰の枝がある.明るいところでさかんに光合成生産をおこなっている枝と,光不足で困窮している枝とがあるとき,そのあいだで有機物の流れはあるだろうか?余っているところから足りないところへ流れていく可能性も考えられるが,実際には逆である.明るいところに優先的に有機物が流れており,日陰の枝は助けてもらえないどころか、むしろ乏しい生産物まで吸い上げられて成長が止る,さらには枯死してしまうという現象が報告されている(STOLL & SCHMID 1998,TAKENAKA 2000, HENRIKSSON 2001).一度日陰になってしまった枝の身になると踏んだり蹴ったりだが,個体全体としては,あまり見返りが期待できない枝を力学的・生理的に支える投資を節約して,その分,日当たりのよいところに資源を集中して光を受ける体制を充実させたほうがメリットは大きそうだ.では,どのような場合にどれだけのメリットがあるのか.定量的に評価しようとすると,部分から全体への統合,そして構造作りのコストの評価が不可欠になるが,これらは今後の課題である.
林の地面近くには,高木の日陰に甘んじながら暮らしている木々がいる.そのなかには,いくら環境に恵まれても数メートル以上にはならない低木樹種もあれば,光さえあれば成長して林冠に到達する高木樹種の稚樹もある.
KOHYAMA (1987)は,屋久島の常緑広葉樹林の林床に生育している木々のなかなら10種を選んで,高さと水平方向のひろがりとを調べた.それらのデータを並べてみると,高さ指向の樹種と、横方向の広がり指向の樹種とにグループ分けすることができた.KOHYAMAは,それぞれのグループの構造的な特徴は林のなかでの木の生き方と関連しているのではないかと考えた.細長く上に伸びる高さ指向の種類では、いつの日か上の木が死んで明るくなったときの高さ競争に勝って,いちはやく林冠を埋める高木となることを目指す生き方をしており,いっぽう横に広がる種類は、今の光不足の環境のなかで、上からくる光をすこしでも多く受けて長生きしようという生き方をしているのではないかという仮説である(図5).
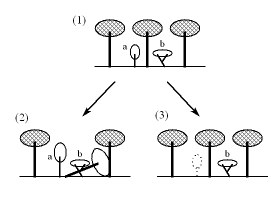
| 図5 光不足の林のなかで生きる木の構造の2タイプ(1).aタイプは広がりは少ないがその分高かく,上の木が死んで明るくなるといちはやく成長してその空隙を埋める(2).bタイプは高さよりも広がりに重点をおいた構造をしており,長期間の光不足のもとでも生存できる(3). |
この議論の前提となっているのは,高さと広がりとのトレードオフ関係(どちらかを優先すると他方が犠牲になる関係)である.枝を上に伸ばすにしろ横に伸ばすにしろ,そのための材料が必要だし,既存部分も伸びた枝を支えられるように補強する必要がある.限られた有機物をもとでに形作りをするのだから,高さと広がりがトレードオフの関係にあることは明らかである.その定量的な関係は,多数の測定データから経験的に推定することはできる.しかし,力学的強度の計算にもとづいて必要なコストを見積もり,植物がしたがわざるを得ない制約を理詰めで示すことができれば,より深い理解が得られるはずである.ここでも,幹や枝のコストを定量定期に評価することが重要なポイントとなってくる.
ところで,KOHYAMAの仮説のひとつの特徴は,一時点の受光効率だけではなく,将来の成功を視野において構造の意味を考えている点である.成長することで資源を探す植物では,現在の構造は過去の探索の歴史を反映しているとともに,将来の探索の足場でもある.構造作りのコストと利得を生涯にわたって積分して評価することできれば,構造と生き方の関係を理解はより深まるだろう.時間断面から生涯への定量的な積分もこれから特に注目するべき方向だろう.
枝や幹は先端に近いほど細く、基部ほど太いのがふつうである(図6).どこも同じ太さだったり, 逆に末端ほど太くなっていたら,とても奇妙に見えるだろう.各部の太さをくわしく調べてみると,1本の枝が2本に枝分かれしているところでは,枝分かれ前の枝の断面積と,枝分かれ後の2本の枝の断面積を合計したものがほぼ等しいという関係がみつかる.さらに,その断面積は,そこから先の枝についている葉の総量と比例関係にあることが多い.このパターンの一般性については本特集の城田・作田両氏の記事でくわしく述べられている.

| 図6 落葉期のミズナラ. |
SHINOZAKI et al. (1964)は,この現象にもとづいて“パイプモデル”を提唱した.このモデルでは,一枚一枚の葉にそれを力学的・生理的に支えるパイプが連なったものが体をつくる単位で,これを束ねてところどころ枝分かれさせたものが植物体だと見る.これは上のパターンをスマートに説明できるモデルだが、単位パイプそのものは実体をともなうものではない.また、力学的・生理的に支えるのに必要な太さのパイプといっても、何のためにどれだけの太さが必要なのかは示されてない。
そもそも力学的支持に必要な太さと,水の供給など生理的な支持に必要な太さが同じである必然性はないので,「力学的・生理的に支えるのに必要な太さ」という表現は不正確だ.実際には2つの要求のうちの大きいほうに合わせた太さを持っていて,もうひとつの要求にたいしては過剰にこたえているのだろう.本特集中の種子田・舘野両氏の記事は,針葉樹など仮導管しか持たないものでは生理的支持の要求が大きくて力学的には余裕がり,多くの広葉樹のように導管を持つものでは力学的支持の要求のほうが大きくて通導機能に関しては余裕がある,という興味深い結果を紹介している.枝の太さのパターンを記述するのではなく,木の生存のためにどれだけの太さが必要なのかを明らかにする研究は,木の構造と機能の理解を深めるうえできわめて重要な意味を持つ.
ここまでしばしば枝や幹の太さの成長について言及してきた,枝や幹が太さを持つからこそ,空間内の資源獲得はコストとの兼ね合いのなかでなされることになる.さまざまなサイズの木の測定データから,ある高さや広がりを持つ木はこれだけの量の枝や幹を必要とするのだと経験的に決めることはできる.これにもとづいてコストと利得のかねあいを考えることもできる.しかし,ある構造体が自重や風圧などの外力にたいしてある安全率で対抗するために必要な太さは,材料力学の分野の知識を使って計算することが可能だ.有限要素法というコンピュータの計算力に依存して力づくで解析してしまう手段なら、樹木の複雑な構造も扱える(MATTHECK, 1998).このような手法を使った生態学的な研究の例はまだごく少ないが,積極的に活用すれば得られるものは大きいはずである.
なお,力学的な支持に必要な太さは,その場の風の強さや降雨,降雪にも依存するだろう. また,生理的要求,とくに水の通導能力も,物理環境や葉の性質によって蒸散量が大きく変化することも考えて評価する必要がある.さらに,どれだけ安全率を見込んだ力学的強度とするかは,植物の生き方と関連するだろう.隣接個体との競争な熾烈な場合には,安全率を多少犠牲にしても高く伸びたほうが成功の確率が高いかもしれない.一本の木だけを見るのではなくその置かれた環境全体も考慮しなくてはいけない.
樹木の構造にみられるパターンをいくつか紹介しながら,その意義を評価するためにどのようなアプローチが必要かを考えてきた.その中で,部分から全体へ,そして定性的議論から定量的議論へという方向性の重要性を指摘した.枝レベルから個体レベルへのスケールアップや時間的な積分,トレードオフ関係の定量的な評価,外部環境との相互作用などがとくに重要だろう.
複数の要素の知識を統合し,個体レベルへとスケールアップし,あるいは時間積分を行って定量的な議論をするためには,コンピュータを使った数値シミュレーションが必要な場面が多くなる.この特集のなかでも, 梅木氏はシラカンバの樹形形成をモデル化した例を紹介している.コンピュータ中に植物を再構成してその構造や機能を解析する研究は,FISHER & HONDA (1977)を先がけとして,現在も進められている(たとえばPEARCY & YANG 1996, NIKLAS 1999, MURAOKA et al.. 1998, TAKENAKA et al. 2001)が,もっと広く使われてしかるべき研究方法だろう.
なお,本稿では地上部にのみ注目して話を進めてきた.特集の他の記事でも,樹木の地下部に触れたものはない.地下部の構造と機能の研究の難しさは,地上部のそれに輪をいくつもかけたものである.地上からは根や土壌中の環境は見えない.見ようとすると構造を乱す.そんな方法論上の困難が研究の大きな障害になっており,地上部に比べて研究は遅れている.しかし,地下部を無視したままではいつになっても全体の理解には至らない.地下部の構造と機能の研究は,隠れたフロンティアと言えよう.
本稿では,また,シュートの生成,伸長,肥大,枯死の生理的メカニズムなどにも触れていない.これらについても興味深い研究課題はたくさんある.樹木の形態形成パターンの研究,そのパターンの背景にある生理的メカニズムをさぐる研究,そして資源をもとめて競争しながら生きている樹木にとっての形の意味の研究.これらの研究が互いを意識しつつ発展するその先に,樹木の生き方のより深い理解が待っているのだろう.
FISHER, J.B. & HONDA, H. 1977 Bot. Gaz. 138:377-384.
FORD, E.D. et al. 1990 J. theor. Biol. 146:15-36.
HENRIKSSON, J. 2001 Oecologia 126:482-486.
HONDA, H. 1971 J. theor. Biol. 31:331-338.
KOHYAMA, T. 1987 Funct. Ecol. 1:399-404.
MATTHECK, C. 1998 Design in nature: Learning from trees. Springer.
MURAOKA, H. et al. 1998 Ann. Bot. 82:297-307.
NIKLAS, K.J. 1999 J. Exper. Bot. 50:39-52.
PEARCY, R.W. & YANG, W. 1996 Oecologia 108:1-12.
SHINOZAKI, K. et al. 1964 Jap. J. Ecol. 14:97-105.
SPRUGEL, D.G. 1991 Annu. Rev. Ecol. Syst. 22: 309-334.
STOLL, P. & SCHMID, B. 1998 J. Ecol. 86:934-945.
TAKENAKA, A. 1994 J. Plant Research 107:321-330.
TAKENAKA, A. 1997 Tree Physiology 17:205-210.
TAKENAKA, A. 2000 Tree Physiol. 20:987-991.
TAKENAKA, A. et al. 2001 Funct. Ecol. 15:660-668
YAGI, T. & KIKUZAWA, K. 1999 J. Plant Research 112:343-352.